Na transsectie van de perifere zenuw van de bovenste ledematen en chirurgisch herstel, herwinnen sommige patiënten een goede sensomotorische functie, terwijl andere dat niet doen. Het begrijpen van perifere en centrale mechanismen die bijdragen aan herstel kan de ontwikkeling van nieuwe therapeutische interventies vergemakkelijken. Plasticiteit na perifere zenuwdoorsnijding is aangetoond door de neuroaxis in diermodellen van zenuwbeschadiging. De hersenveranderingen die optreden na perifere zenuwtranssectie en chirurgisch herstel bij mensen zijn echter niet onderzocht. Bovendien is de mate waarin regeneratie van perifere zenuwen functionele en structurele hersenveranderingen beïnvloedt, niet gekarakteriseerd. Daarom vroegen we of functionele veranderingen gepaard gaan met structurele veranderingen in de grijze en/of witte stof en of deze veranderingen verband houden met sensorisch herstel? Om deze belangrijke problemen aan te pakken hebben we (i) de regeneratie van perifere zenuwen beoordeeld; (ii) gemeten functionele magnetische resonantie beeldvorming hersenactivatie (bloedzuurstofniveau-afhankelijk signaal; BOLD) in reactie op een vibrotactiele stimulus; (iii) de structurele hersenplasticiteit van grijze en witte stof onderzocht; en (iv) gecorreleerde sensorische herstelmaatregelen met veranderingen in grijze stof bij perifere zenuwtranssectie en chirurgische reparatiepatiënten. Vergeleken met de gezonde contralesionale zenuw van elke patiënt, hebben doorgesneden zenuwen 1.5 jaar na transsectie en herstel een verminderde zenuwgeleiding, geleidend met verminderde amplitude en verhoogde latentie. Vergeleken met gezonde controles hadden patiënten met perifere zenuwtranssectie en chirurgisch herstel een veranderde bloedzuurstofniveau-afhankelijke signaalactiviteit in de contralesionale primaire en secundaire somatosensorische cortex, en in een reeks hersengebieden die bekend staan als het 'taakpositieve netwerk'. Bovendien werden verlagingen van grijze stof vastgesteld in verschillende hersengebieden, waaronder de contralesionale primaire en secundaire somatosensorische cortex, in dezelfde gebieden waar bloedzuurstofafhankelijke signaalverlagingen werden vastgesteld. Bovendien was het dunner worden van grijze stof in de post-centrale gyrus negatief gecorreleerd met metingen van sensorisch herstel (mechanische en trillingsdetectie) die een duidelijk verband aantonen tussen functie en structuur. Ten slotte identificeerden we verminderde fractionele anisotropie van witte stof in de rechter insula in een regio die ook verminderde grijze stof vertoonde. Deze resultaten geven inzicht in hersenplasticiteit en structuur-functie-gedragsrelaties na zenuwbeschadiging en hebben belangrijke therapeutische implicaties.
Sleutelwoorden: corticale dikte; fMRI; diffusie tensor beeldvorming; plasticiteit; perifere zenuwbeschadiging
Afkortingen: BA=Brodmann-gebied; BOLD = afhankelijk van het zuurstofgehalte in het bloed; fMRI=functionele magnetische resonantie beeldvorming;
PNIr=perifere zenuwtranssectie en chirurgisch herstel; S1=primaire somatosensorische cortex; S2=secundaire somatosensorische cortex
Inhoud
Introductie
Na transsectie van de perifere zenuw van de bovenste ledematen en chirurgisch herstel (PNIr) is ongeveer 25% van de patiënten 1.5 jaar na de operatie niet weer aan het werk gegaan (Jaquet et al., 2001). Bovendien is ? 57% van de patiënten met zenuwletsel tussen de 16-35 jaar (McAllister et al., 1996); dus kan een lange levensduur van invaliditeit en economische moeilijkheden gepaard gaan met transsectie van de zenuwen van de bovenste ledematen. Het begrijpen van de centrale en perifere vertakkingen van perifere zenuwbeschadiging kan de ontwikkeling van nieuwe therapeutische strategieën en interventieprogramma's vergemakkelijken.
Het is niet bekend hoe de hersenen reageren op PNIr bij mensen. Dierstudies hebben echter aangetoond dat plasticiteit in de somatosensorische cortex onmiddellijk begint na perifere zenuwtranssectie, en dat corticale kaarten 1 jaar na volledige zenuwtranssectie en chirurgisch herstel fragmentarische, niet-continue representaties van de doorgesneden en aangrenzende zenuwen bevatten (Wall et al. ., 1986). Men denkt dat de mechanismen die functionele plasticiteit vergemakkelijken, de onmiddellijke ontmaskering van reeds bestaande projecties van aangrenzende corticale en subcorticale niveaus omvatten, en het op lange termijn ontspruiten van axonen op meerdere niveaus van de neuro-as, inclusief de primaire somatosensorische cortex (S1) (Florence en Kaas , 1995; Hickmott en Steen, 2005).
Beeldvormingsonderzoeken van de menselijke hersenen hebben de bevindingen van diermodellen bevestigd met de identificatie van veranderde functionele MRI-activeringskaarten als gevolg van ruggenmergletsel, amputatie, teen-naar-duimoverdracht en bij patiënten met carpeltunnelsyndroom (Lotze et al., 2001; Manduch et al., 2002; Jurkiewicz et al., 2006; Napadow et al., 2006). Bovendien hebben structurele MRI-onderzoeken recentelijk veranderingen in grijze en witte stof gevisualiseerd na traumatische verwondingen en in diverse pathologische aandoeningen, waaronder amputatie van ledematen en chronische pijn (Apkarian et al., 2004; Draganski et al., 2006; Davis et al., 2008; Geha et al., 2008; mei 2008). Aangenomen wordt dat veranderingen in grijze stof verband houden met veranderingen in celgrootte, atrofie en/of verlies van neuronen of glia, terwijl veranderingen in witte stof worden beïnvloed door axonale degeneratie en verlies van myeline (Beaulieu, 2002; May, 2008).
Een krachtige benadering om mechanismen van pathologie en plasticiteit af te bakenen is om functionele en structurele beeldvormingstechnieken voor grijze en witte stof te combineren. We hebben eerder gemeld dat patiënten met volledige PNIr van de bovenste ledematen ernstige somatosensorische tekorten behielden die 41.5 jaar na de operatie aanhouden (Taylor et al., 2008a). Op basis van deze bevindingen redeneerden we dat deze patiënten functionele en structurele hersenveranderingen zouden vertonen in belangrijke somatosensorische hersengebieden. Daarom veronderstelden we in deze studie dat PNIr-patiënten zouden hebben: (i) verminderde bloedzuurstofniveau-afhankelijke (BOLD) reacties op vibrerende stimulatie van het doorgesneden zenuwgebied, in het gebied van S1 dat het gewonde bovenste lidmaat vertegenwoordigt en in de secundaire somatosensorische cortex (S2); (ii) een overeenkomstige vermindering van de corticale dikte in deze gebieden van de contralesionale S1 en S2; (iii) een correlatie tussen veranderingen in corticale dikte en psychofysische metingen van somatosensorische functie (trillings- en aanrakingsdetectiedrempels); en (iv) verminderde fractionele anisotropie (een maat voor de integriteit van de witte stof) in witte stof die in/uit deze somatosensorische corticale gebieden stroomt.
Methoden
vakken
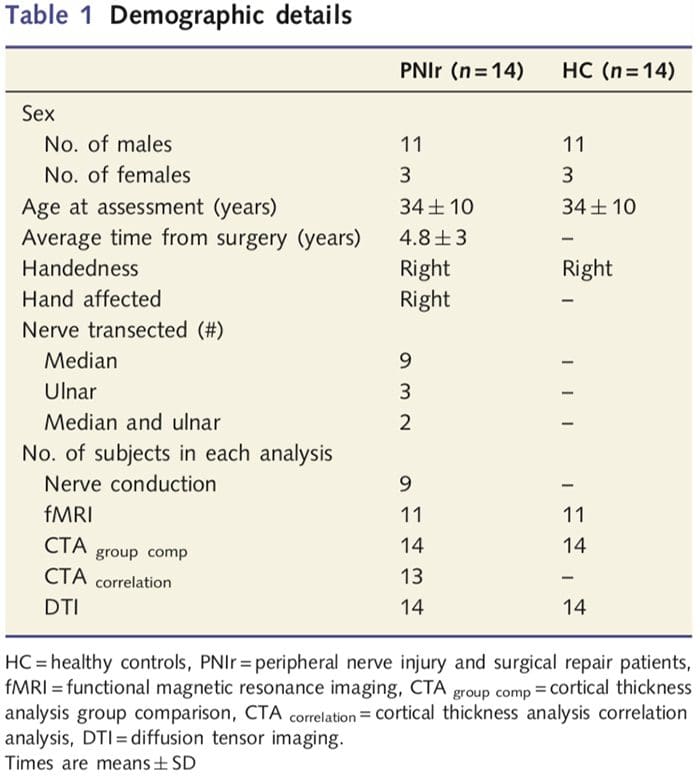
We rekruteerden 27 patiënten met een volledige transsectie van de mediane en/of nervus ulnaris gevolgd door chirurgisch herstel van plastisch chirurgen verbonden aan het University of Toronto Hand Program tussen juni 2006 en mei 2008. Van dit grotere cohort werden 14 pijnvrije patiënten (drie vrouwen , 11 mannen; 34 - 10 jaar) met volledige doorsnijding van de rechter mediane en/of nervus ulnaris werden in het onderzoek geïncludeerd [om verwarring in verband met de aanwezigheid van pijn en lateraliteitspatiënten met pijn (n=6) en linkszijdige laesies te voorkomen (n=7) werden uitgesloten van deze analyse]. Alle patiënten ondergingen microchirurgische zenuwreparatie ten minste 1.5 jaar voorafgaand aan deelname aan de studie (hersteltijd varieerde van 1.5 tot 8 jaar). Daarnaast rekruteerden we 14 gezonde controles op leeftijd en geslacht (3 vrouwen, 11 mannen; 34 - 10 jaar). Alle proefpersonen gaven geïnformeerde schriftelijke toestemming voor procedures die waren goedgekeurd door de University Health Network Research Ethics Board. Alle proefpersonen waren rechtshandig (bepaald met behulp van de Edinburgh handigheidsinventaris: Oldfield, 1971) en hadden geen voorgeschiedenis van neurologisch letsel of chronische pijn (voor of na zenuwdoorsnijding). Zie Tabel 1 voor demografische details.
studie ontwerp
Alle proefpersonen namen deel aan een beeldvormingssessie die omvatte: (i) functionele magnetische resonantiebeeldvorming (fMRI) als reactie op vibrotactiele stimuli toegepast op de rechter wijsvinger (binnen het mediane zenuwgebied); (ii) een anatomische scan met hoge resolutie van de hele hersenen, verkregen voor beeldregistratie en voor de analyse van corticale grijze stof; en (iii) twee diffusie tensor-beeldvormingsscans voor de beoordeling van de integriteit van de witte stof. Voorafgaand aan de beeldvorming werden de proefpersonen geïnstrueerd in het basisontwerp van het experiment en eraan herinnerd om zo stil mogelijk te blijven gedurende de duur van de scan.
De proefpersonen waren vrij om zich op elk moment uit het onderzoek terug te trekken. Daarnaast werd voor alle proefpersonen een sensorische en motorische beoordeling uitgevoerd (Taylor et al., 2008a). Aangezien de drempels voor detectie van aanraking en trillingen gecorreleerd waren met de corticale dikte, is een beschrijving van deze methoden hieronder opgenomen (andere psychofysische maatregelen zullen elders worden gerapporteerd).
Trillingsdrempel
Trillingsdetectiedrempels werden bepaald met behulp van een draagbare Bio-Thesiometer (Bio-Medical Instrument Company, VS). Het apparaat heeft een sonde van 12 mm die op de distale falanx van de rechter wijsvinger (D2) is geplaatst. Drempels werden bepaald met behulp van de methode van limieten: de amplitude (spanning) werd geleidelijk verhoogd totdat het onderwerp aangaf dat ze de stimulus waarnamen. Trillingsdrempels werden driemaal verkregen en een gemiddelde waarde werd berekend. Tijdens het testen van de trillingsdrempel kregen de proefpersonen de opdracht om hun ogen te sluiten en de rug van hun hand op een ondersteunend kussen te laten rusten.
Mechanische detectiedrempel
Mechanische detectiedrempels werden bepaald met behulp van een gestandaardiseerde set von Frey-filamenten (OptiHair2 Marstock Nervtest, Duitsland) die 12 logaritmisch op afstand van elkaar gekalibreerde filamenten bevatten die krachten leverden van 0.25, 512-12 mN. De diameter van het contactoppervlak van alle 0.4 filamenten was ~2 mm. De proeven werden uitgevoerd met de ogen van de proefpersoon gesloten en de handen op een zacht kussen. Sondes werden in een oplopende reeks aangebracht en proefpersonen moesten elke keer dat ze een sonde de rechter DXNUMX-vingertop voelden raken, een reactie geven. Dit proces werd drie keer herhaald. De kracht voor het filament die in ten minste twee van de drie onderzoeken werd gedetecteerd, werd gerapporteerd als de mechanische detectiedrempel van die proefpersoon.
Zenuwgeleiding testen
Patiënten namen deel aan bilaterale onderzoeken naar sensorische en motorische zenuwgeleiding in de elektromyografie (EMG) kliniek van het Toronto Western Hospital. Voor motorische zenuwgeleiding werd de stimulerende elektrode bij de pols en elleboog (afzonderlijk) geplaatst en de opname-elektrode werd over de abductor pollicis brevis geplaatst, voor beoordeling van de medianuszenuw, of de abductor digiti minimi voor beoordeling van de nervus ulnaris. Voor het testen van de sensorische zenuwen werd de opname-elektrode op de pols geplaatst en de stimulerende elektrode op de cijfers D2, D3 en D5. Een senior, ervaren neuroloog van de Toronto Western Hospital EMG Clinic (Dr. Peter Ashby) beoordeelde alle klinische beoordelingen om te bepalen welke zenuwen normale/abnormale reacties vertoonden. Aangezien bekend is dat amplitude- en latentiemetingen aanzienlijk variëren tussen proefpersonen (vanwege factoren zoals de dichtheid van innervatie, de diepte van de zenuw en de dikte van de huid van een individuele proefpersoon) (Kimura, 2001) is de niet-doorgesneden zenuw van elke patiënt dienden als hun eigen controle voor vergelijking met waarden van de doorgesneden zijde. Bij die patiënten met detecteerbare zenuwgeleidingsreacties werden gepaarde t-tests uitgevoerd om het verschil in latentie- of amplitudemetingen tussen de doorgesneden en contralesionale niet-doorgesneden zenuwen van elke patiënt te beoordelen.
Beeldparameters:
Hersenbeeldgegevens werden verkregen met behulp van een 3T GE MRI-systeem uitgerust met een achtkanaals phased array-kopspoel. De proefpersonen werden in rugligging op de MRI-tafel gelegd en het hoofd van elke proefpersoon werd opgevuld om beweging te verminderen. Full-brain fMRI-gegevens werden verkregen met behulp van echo vlakke beeldvorming (28 axiale plakjes, gezichtsveld (FOV) = 20 x 20 cm, 64 x 64 matrix, 3.125 x 3.125 x 4 mm voxels, echotijd (TE) = 30 ms, herhaling tijd (TR) = 2000 ms). De scantijd was 5 min en 8 s (154 frames). Tijdens het scannen werd een niet-pijnlijke vibrotactiele stimulus van 12 Hz toegepast op de distale falanx van de rechter D2 met behulp van ballonmembranen aangedreven door perslucht (apparaat vervaardigd door Dr. Christo Pantev; www.biomag.uni-muenster.de). Stimuli werden toegediend in blokken van 10 seconden, afgewisseld met 20 seconden rust, voor een totaal van 10 stimulatieblokken en 10 rustblokken. De eerste 8 s (4 TR's) aan gegevens die van elke run werden verkregen, werden weggegooid om fMRI-signaalevenwicht mogelijk te maken. De proefpersonen kregen de opdracht om hun ogen tijdens het scannen gesloten te houden en zich op de stimuli te concentreren. Een driedimensionale (3D) anatomische scan met hoge resolutie van het hele brein (124 sagittale plakjes, 24 x 24 cm FOV, 256 x 256 matrix, 1.5 x 0.94 x 0.94 mm voxels) werd verkregen met een T1-gewogen 3D verwende gradiëntechosequentie (één signaalgemiddelde, kantelhoek = 20?, TE ?5 ms). Daarnaast werden twee diffusie tensor imaging scans (38 axiale plakjes, FOV 24 x 24 cm, 128 x 128 matrix, 1.875 x 1.875 x 3 mm voxels) verkregen langs 23 richtingen met een b-waarde van 1000smm2. Elke run bevatte ook twee volumes zonder diffusieweging.
fMRI-analyse
Gegevens werden geanalyseerd met Brainvoyager QX v1.8 (Brain Innovaton, Maastricht, Nederland). Voorbewerking omvatte: 3D-bewegingscorrectie, slice-scantijdcorrectie, lineaire trendverwijdering, hoogdoorlaatfiltering (vijf cycli per run) en ruimtelijke afvlakking met een Gauss-kernel van 6 mm volledige breedte op half maximum (FWHM). fMRI-gegevenssets werden geïnterpoleerd naar voxels van 3 x 3 x 3 mm, geregistreerd op het anatomische beeld met hoge resolutie en genormaliseerd naar de standaard Talairach-ruimte (Talairach en Tournoux, 1988). Voxels worden gerapporteerd als 1 x 1 x 1 mm. Gegevens werden geanalyseerd met behulp van het algemene lineaire model; het model werd verkregen door de functie van de boxcar van het tijdsverloop van tactiele stimulatie te convolueren met de standaard hemodynamische responsfunctie. Om groepsverschillen in activeringspatronen te identificeren, werd een fixed effects-analyse uitgevoerd met de contrasten: (i) gezonde controles: stimulatie 4 rust; (ii) PNIr: stimulatie 4 rust; en (iii) gezonde controles 4 PNIr. Activeringskaarten werden gedrempeld op een gecorrigeerde waarde van P50.05 (afgeleid van een niet-gecorrigeerde P50.0001 en 120 mm3 aaneengesloten voxels zoals eerder gerapporteerd: Taylor en Davis, 2009); dit werd ook gevalideerd door een Monte Carlo-simulatie uit te voeren met de AlphaSim-toepassing die is geïmplementeerd in de analyse van Functional Neuroimage (AFNI)-software. Deze analyse omvatte alleen de 11 patiënten die aanhoudende transsectie van de rechter medianuszenuw (n=9) of de rechter mediaan- en ulnaire zenuw (n = 2) (dwz de drie patiënten met een zuivere rechter ulnaire zenuwdoorsnijding werden niet opgenomen in deze analyse ).
Corticale dikte-analyse
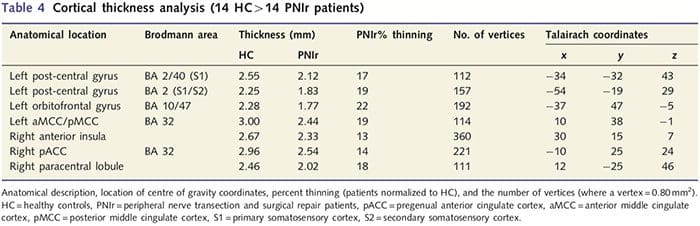
Corticale dikte-analyse werd uitgevoerd met behulp van Freesurfer (//surfer.nmr.mgh.harvard.edu); methoden zijn elders in detail beschreven (Dale et al., 1999; Fischl et al., 1999a, b; Fischl en Dale 2000). In het kort werden T1-gewogen anatomische datasets met hoge resolutie geregistreerd in de Talairach-atlas (Talairach en Tournoux, 1988). Dit werd gevolgd door normalisatie van de intensiteit, het strippen van de schedel en scheiding van de hemisferen. Vervolgens werden de grenzen van de wit/grijze stof (het witte oppervlak genoemd) en de grijs/CSF (het pialoppervlak genoemd) geïdentificeerd en gesegmenteerd. De afstand tussen de witte en pial-oppervlakken werd vervolgens berekend op elk punt in elk halfrond van de hersenen. Om groepsverschillen tussen de 14 patiënten en 14 leeftijd/geslacht-gematchte controles te identificeren, werd een algemene lineaire modelanalyse uitgevoerd op elk punt in de hersenen. Omdat de corticale topografie van het individu inherent heterogeen is, werd voorafgaand aan de statistische analyse een 5 mm FWHM ruimtelijke afvlakkingskernel toegepast. Gegevens worden weergegeven op een gecorrigeerde P50.05 (afgeleid van een niet-gecorrigeerde P50.0075 en 102 aangrenzende hoekpunten); dit werd berekend door een Monte Carlo-simulatie uit te voeren met AlphaSim. Een hoekpunt vertegenwoordigt een punt op een tweedimensionale plaat, en in deze studie is de afstand tussen twee hoekpunten 0.80 mm2.
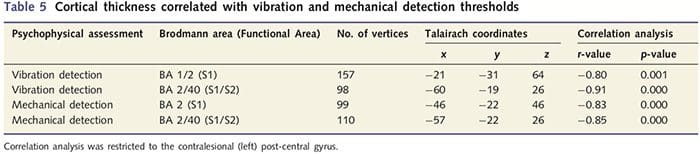
Omdat patiënten significante tekortkomingen vertoonden in somatosensorische functie binnen het doorgesneden zenuwgebied, veronderstelden we dat metingen van somatosensorische functie (vibratie en aanrakingsdetectie) zouden correleren met corticale dikte in de contralesionale post-centrale gyrus (primaire en secundaire somatosensorische cortex). Daarom hebben we correlatieanalyses uitgevoerd in de patiëntengroep tussen: (i) corticale dikte en trillingsdetectiedrempel; en (ii) corticale dikte en aanraakdetectiedrempels. Eén patiënt heeft de psychofysische beoordeling niet voltooid; daarom omvatte deze analyse 13 PNIr-patiënten. Om te bepalen of er een verband was tussen corticale dikte en hersteltijd, werd bovendien ook een correlatieanalyse tussen deze twee metingen uitgevoerd. Deze correlatieanalyses waren beperkt tot de contralesionale post-centrale gyrus door een masker (overgenomen uit de ingebouwde atlas van Freesurfer) in het algemene lineaire model op te nemen. Er werd een Monte Carlo-simulatie uitgevoerd die beperkt was tot het aantal hoekpunten binnen contralesionale post-centrale gyrus; afbeeldingen worden weergegeven met een gecorrigeerde P50.05 (afgeleid van een niet-gecorrigeerde P50.0075 en 68 aangrenzende hoekpunten).
Diffusie Tensor Imaging-analyse
Diffusietensorbeeldverwerking werd uitgevoerd met DTiStudio (www.MriStudio.org) en FSLv.4.0 (www.fmrib.ox.ac.uk/fsl/). Afbeeldingen werden eerst opnieuw uitgelijnd met de automatische beeldregistratietool die in DTiStudio is geïmplementeerd, waarbij de eerste B0-afbeelding in de eerste reeks als sjabloon werd gebruikt. Dit proces corrigeert voor beweging van het onderwerp en wervelstroomvervorming. Alle afbeeldingen werden vervolgens visueel geïnspecteerd om de beeldkwaliteit en de uitlijning van de afzonderlijke diffusietensorbeeldvormingsruns te beoordelen. Als een artefact werd gedetecteerd, werd het plakje verwijderd voordat het gemiddelde van de twee afzonderlijke diffusietensorbeeldvormingsruns werd berekend. Individuele FA-kaarten werden berekend met behulp van de DTIFIT-tool die in FSL is geïmplementeerd. Voxel-gewijze statistische analyse werd uitgevoerd om groepsverschillen in de gemiddelde fractionele anisotropie te identificeren met behulp van Tract Based Spatial Statistics; voor een volledige beschrijving van deze methoden zie Smith et al. (2006). Kort gezegd, afbeeldingen werden niet-lineair geregistreerd op een doelafbeelding (MNI152), de gemiddelde afbeelding werd vervolgens gemaakt van alle datasets en deze afbeelding werd vervolgens uitgedund om alle traktaten weer te geven die voor alle onderwerpen gemeenschappelijk waren. De hoogste fractionele anisotropiewaarden van elke proefpersoon werden vervolgens op het skelet geprojecteerd door te zoeken in witte stof loodrecht op elk punt op het skelet van witte stof. Vervolgens werd een voxel-gewijze statistische analyse van de hele hersenen uitgevoerd tussen groepen (14PNIr en 14 gezonde controles) en beelden werden gecorrigeerd voor de hele hersenen op P50.05. Bovendien werd een analyse van het interessegebied uitgevoerd in witte stof traktaten grenzend aan de contralaterale S1, thalamus en bilaterale anterieure en posterieure insula. Deze regio's zijn gekozen omdat ze eerder betrokken waren bij aspecten van somatosensatie en omdat ze overeenkomen met regio's die werden geïdentificeerd in de fMRI- en corticale dikte-analyse (CTA) groepsanalyses. Van belang zijnde gebieden werden als volgt op het skelet van de witte stof getekend: (i) Het contralaterale S1-gebied van belang ontstond mediaal op de kruising tussen het skelet van de witte stof van de corona-radiata en het skeletgedeelte dat de post-centrale gyrus invoerde; eindigend aan het einde van het kanaal binnen een bepaalde plak.
In de z-richting strekte het interessegebied zich uit van z=49 tot 57; witte stof traktaten die het handgebied bevoorraden. (ii) Het van belang zijnde contralaterale thalamusgebied was beperkt tot sporen van witte stof rond de achterste en mediale thalamische kernen (kernen die betrokken zijn bij de somatosensorische functie), zich uitstrekkend van z = -1 tot 4. (iii) Insulaire gebieden van belang werden bilateraal binnen witte stof grenzend aan de voorste en achterste insula op basis van eerder door ons laboratorium gepubliceerde criteria (Taylor et al., 2008b). Het interessegebied strekte zich uit van z = 2 tot 8. Fractionele anisotropiewaarden werden geëxtraheerd uit elk van deze interessegebieden en een multivariate variantieanalyse (MANOVA) werd uitgevoerd met behulp van het Statistical Package for the Social Sciences v13.0 (SPSS Inc, Chicago), die fractionele anisotropiewaarden bevatte voor alle zes interessegebieden.
Resultaten
Tabel 1 geeft demografische details voor studiedeelnemers. Alle 14 patiënten hadden een volledige transsectie van de rechter mediane en/of nervus ulnaris gevolgd door microchirurgische reparatie ten minste 1.5 jaar voorafgaand aan deelname aan het onderzoek. De tijd van operatie tot testen varieerde van 1.5 tot 8 jaar met een gemiddelde (?SD) van 4.8 ? 3 jaar. Patiënten en controles niet (34 - 10 jaar beide groepen; t = 0.04; P = 0.97).
Psychofysica
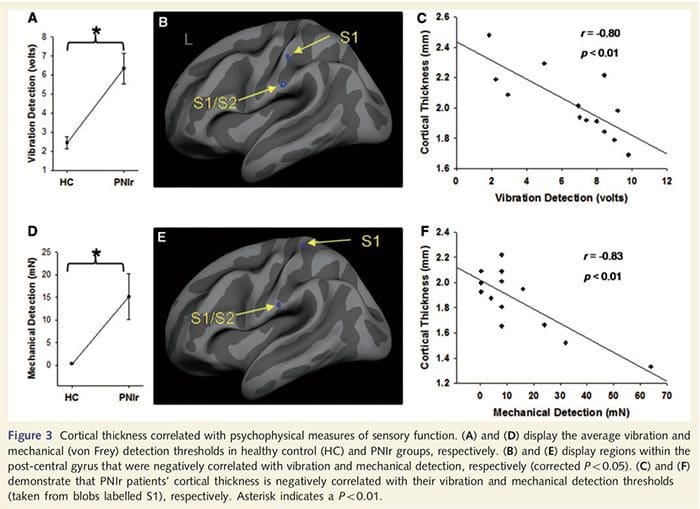
Trillingsdrempels werden berekend uit alle drie de metingen, aangezien eenzijdige variantieanalyse (ANOVA) geen significante verschillen aangaf tussen de drie onderzoeken [F (25, 1)=0.227, P=0.64]. Trillingen en mechanische detectiedrempels waren significant verminderd bij PNIr-patiënten in vergelijking met gezonde controles (vibratie: t = 4.77, P50.001, Fig. 3A; mechanisch: t=3.10, P=0.005, Fig. 3D).
Zenuwgeleiding testen
Amplitude- en latentiemetingen verkregen van de contralesionale zenuwen van elke patiënt werden door een ervaren neuroloog in de EMG-kliniek van het Toronto Western Hospital als normaal geclassificeerd. Negen van de 14 patiënten voltooiden de zenuwgeleidingstest. Tabel 2 geeft de gemiddelde toename/afname latentie- en amplitudegegevens weer voor sensorische zenuwgeleiding van de pols naar de abductor pollicis brevis (mediaan) of abductor digiti minimi (ulnaire) spieren en voor sensorische geleiding van de pols naar D2 (mediaan) en D5 ( ulnaire) vergeleken met de niet-verwonde contralesionale zenuw van elke patiënt. Van de negen hadden zeven patiënten transsecties die de medianuszenuw omvatten. Van deze zeven had één patiënt geen detecteerbare respons tijdens motortesten en een andere patiënt had geen detecteerbare respons tijdens sensorische tests.
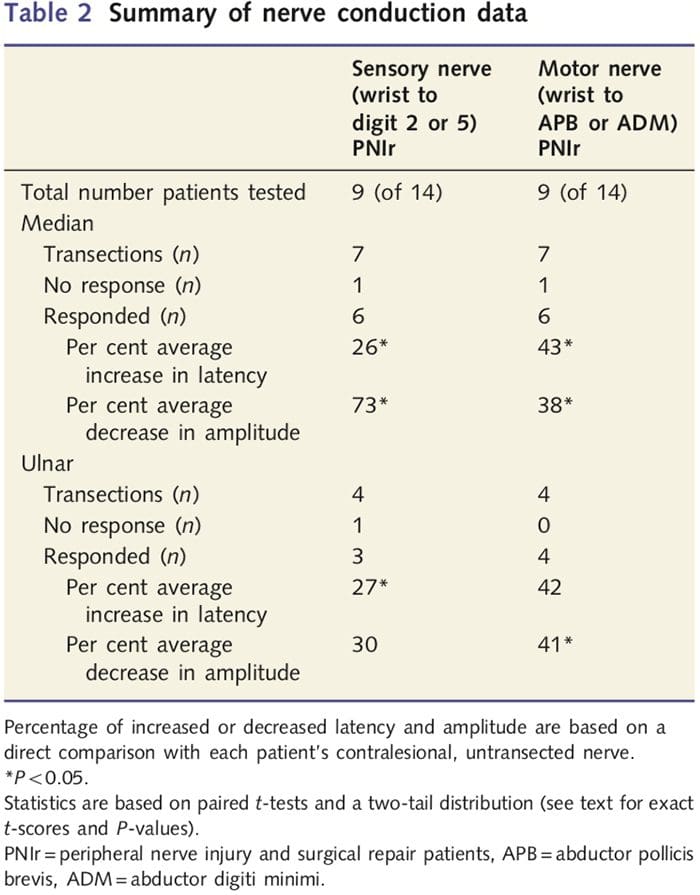 Bij de zes patiënten met detecteerbare responsen waren de motorische geleidingslatenties met 43% verhoogd (t=6.2; P=0.002) en de amplitudes met 38% verlaagd (t=�2.6; P=0.045) wanneer de doorgesneden zenuw van elke patiënt vergeleken met hun niet-geblesseerde kant. Sensorische geleiding in medianuszenuwen onthulde ook een toename van 26% in latentie (t=3.9; P=0.011) en een afname van 73% in amplitude (t=�8.0; P=0.000) vergeleken met normale contralesionale zenuwen. Bij de vier patiënten met ulnaire zenuwtranssecties had één patiënt geen waarneembare respons tijdens het testen van de sensorische zenuwen. Bij die patiënten met reacties waren de motorische latenties van de ulnaire zenuw niet significant verhoogd (t = 2.8; P = 0.070); de amplitudes waren echter aanzienlijk
Bij de zes patiënten met detecteerbare responsen waren de motorische geleidingslatenties met 43% verhoogd (t=6.2; P=0.002) en de amplitudes met 38% verlaagd (t=�2.6; P=0.045) wanneer de doorgesneden zenuw van elke patiënt vergeleken met hun niet-geblesseerde kant. Sensorische geleiding in medianuszenuwen onthulde ook een toename van 26% in latentie (t=3.9; P=0.011) en een afname van 73% in amplitude (t=�8.0; P=0.000) vergeleken met normale contralesionale zenuwen. Bij de vier patiënten met ulnaire zenuwtranssecties had één patiënt geen waarneembare respons tijdens het testen van de sensorische zenuwen. Bij die patiënten met reacties waren de motorische latenties van de ulnaire zenuw niet significant verhoogd (t = 2.8; P = 0.070); de amplitudes waren echter aanzienlijk
afgenomen met 41% (t = -5.9; P = 0.010). Sensorisch testen van de nervus ulnaris toonde een toename van 27% in latentie aan (t = 4.3; P = 0.049) maar geen significante toename in amplitude (t = 3.5; P = 0.072).
Functionele plasticiteit in de primaire somatosensorische cortex
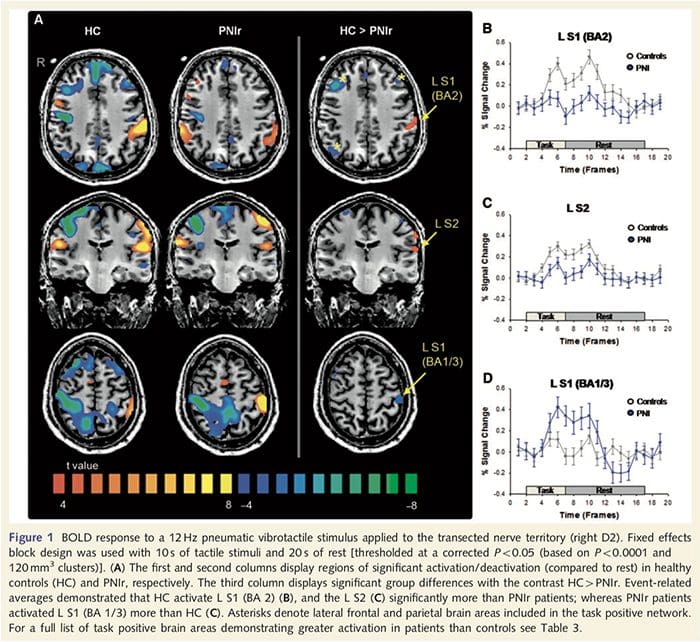
Functionele MRI-kaarten werden berekend uit de 11 PNIr-patiënten met dwarsdoorsneden van de rechter mediane zenuw (patiënten met dwarsdoorsneden van de nervus ulnaris werden uitgesloten van deze analyse) en 11 gezonde controles van dezelfde leeftijd en geslacht. Uit Fig. 1A is het duidelijk dat PNIr-patiënten significant minder activering hebben, vergeleken met gezonde controles, in een regio van S1 die overeenkomt met Brodmann-gebied 2 (BA2) (Talairach en Tournoux, 1988) en S2 (zie tabel 3 voor details) . De gemiddelde gebeurtenisgerelateerde reacties van deze interessegebieden benadrukken de verzwakte BOLD-respons binnen de linker BA2 en linker S2 van de patiënt (respectievelijk Fig. 1B en C). Vreemd genoeg activeerde vibrotactiele stimulatie bij de patiënten een meer superieur deel van de post-centrale gyrus (waarschijnlijk BA1/3) (Talairach en Tournoux, 1988) (Fig. 1A en Tabel 3). Een gebeurtenisgerelateerd gemiddelde (Fig. 1D) toont aan dat gezonde controles minimale activering in dit gebied hadden. Bovendien hadden patiënten significant meer activatie in hersengebieden die gezamenlijk bekend staan als het taakpositieve netwerk (sterretjes in figuur 1). Zie Tabel 3 voor de volledige lijst van geactiveerde taakpositieve hersengebieden. Dit netwerk omvat laterale prefrontale, laterale pariëtale, premotorische en inferieure temporale cortex (Tabel 3): hersengebieden die worden geactiveerd tijdens het uitvoeren van een aandachtsintensieve taak en onderdrukt of inactief zijn tijdens rust of taken die niet cognitief of aandachtig uitdagend zijn (Fox et al. al., 2005; DeLuca et al., 2006; Seminowicz en Davis 2007).
Verminderde grijze stof in de primaire somatosensorische cortex correleert met sensorisch herstel
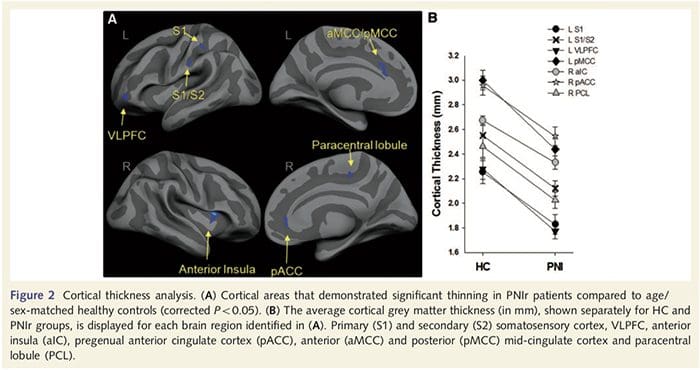
Corticale dikte-analyse bij alle 14 patiënten en 14 gezonde controles van leeftijd/geslacht onthulde verschillende loci van significante corticale verdunning in de PNIr-groep (Fig. 2 en Tabel 4). Specifiek hadden patiënten een vermindering van 13%-22% in corticale dikte in de linker (contralesionale) S1, S2, pregenuale gyrus anterior cingulate, ventrolaterale prefrontale cortex en rechter anterior insula, anterior/posterior mid cingulate gyrus en paracentrale lobule. Interessant is dat de locaties van het dunner worden van grijze stof in de post-centrale gyrus samenvallen met de gebieden met verminderde BOLD na vibrotactiele stimulatie (tabel 4). Omdat we voorkennis hadden van de sensorische tekorten en hersteltijd van de patiënten (dwz de tijd sinds microchirurgische reparatie), vroegen we vervolgens of de corticale dikte van de patiënten in de post-centrale gyrus correleerde met hun sensorische mechanische en trillingsdetectiedrempels, of met hun hersteltijd. Deze analyses lieten een negatieve correlatie zien tussen corticale dikte en trillingsdetectiedrempels in een gebied dat BA1/2 en S2 omvat (P50.001, r=?0.80 en ?0.91, voor BA1/2 en S2 respectievelijk; Fig. 3 en Tabel 5 ). Bovendien waren mechanische detectiedrempels ook negatief gecorreleerd met corticale dikte in een iets meer superieur BA2-gebied en hetzelfde S2-gebied (P50.001, r = -0.83 en -0.85, respectievelijk voor BA2 en S2; Fig. 3 en Tabel 5). We hebben echter geen significante relatie vastgesteld tussen hersteltijd en corticale dikte. Daarom werd corticale dunner worden in de post-centrale gyrus geassocieerd met ernstigere sensorische tekorten. We hebben echter geen significante relatie vastgesteld tussen hersteltijd en corticale dikte. Nogmaals, er was een overeenkomst tussen de corticale verdunning in gebieden die negatief gecorreleerd waren met trillingsstimuli en de regio's die fMRI- en CTA-afwijkingen van de groep vertoonden.
Afwijkingen van de witte stof na zenuwdoorsnijding
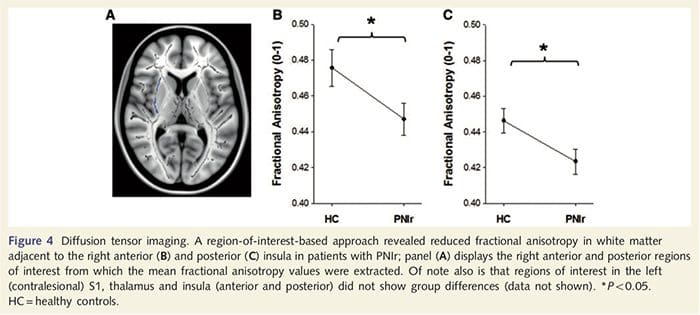
Om de integriteit van de witte stof te beoordelen, hebben we een interessegebiedbenadering gebruikt om verschillen in witte stofgroep te onderzoeken op basis van a priori hypothesen. Regio's van belang waren beperkt tot witte-stofkanalen die de contralesionale S1 en thalamus omringen en voeden. Daarnaast hebben we ook interessegebieden in witte stof getekend naast de linker en rechter, voorste en achterste insula. De insula werd gekozen omdat deze betrokken is bij somatosensorische verwerking en omdat we verminderde grijze stof in het rechter voorste insulair met CTA hebben geïdentificeerd. Deze benadering van interessegebied onthulde dat patiënten significant verminderde fractionele anisotropiewaarden voor witte stof hadden (MANOVA inclusief alle zes interessegebieden) grenzend aan de rechter anterieure [F (1, 26) = 4.39, P = 0.046; Fig. 4A] en posterieure insula [F (1, 26) = 5.55, P = 0.026; Fig. 4B], maar er waren geen groepsverschillen in de witte stof grenzend aan de linker insula (linker anterior insula: P = 0.51; linker posterieure insula: P=0.26), thalamus (P=0.46) of S1 (P=0.46 ).
 Discussie
Discussie
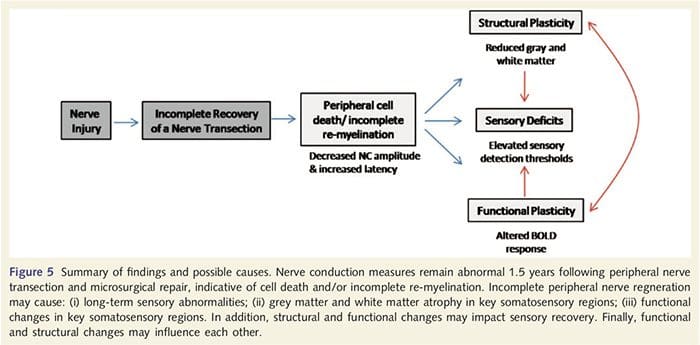
Hier hebben we voor het eerst aangetoond dat er functionele plasticiteit is en structurele afwijkingen in zowel grijze als witte stof in verschillende corticale gebieden na transsectie van de perifere zenuw van de bovenste ledematen en chirurgisch herstel. Deze plasticiteit kan het gevolg zijn van onvolledige regeneratie van perifere zenuwen (perifere celdood en/of onvolledige hermyelinisatie), aangezien zenuwgeleidingsmaatregelen bij deze patiënten ernstige afwijkingen vertoonden. Bovendien tonen onze gegevens aan dat verminderde vibrotactiele-opgewekte fMRI-responsen in de post-centrale gyrus overeenkomen met het dunner worden van grijze stof in de patiëntengroep. Deze resultaten suggereren dat verminderde BOLD-reacties kunnen worden vergemakkelijkt door een vermindering van corticale grijze stof en/of een afname van de afferente input naar de post-centrale gyrus. Bovendien correleerde de corticale dikte binnen dezelfde delen van de post-centrale gyrus negatief met gedragsmetingen van de somatosensorische functie. Dat wil zeggen, verhoogde somatosensorische tekorten waren gecorreleerd met dunnere cortex; beide kunnen te maken hebben met afferente input. Alles bij elkaar genomen suggereren onze gegevens dat onvolledige regeneratie van perifere zenuwen bijdraagt aan somatosensorische stoornissen, atrofie van corticale grijze stof en verminderde fMRI-activering (zie figuur 5 voor een samenvatting van deze bevindingen).
Het is algemeen bekend dat corticale plasticiteit na doorsnijding van perifere zenuwen en chirurgisch herstel door het hele CZS kan optreden bij niet-menselijke primaten (Kaas, 1991). Aangenomen wordt dat deze plasticiteit te wijten is aan het ontmaskeren van voorheen stille synapsen of axonale ontspruiting in gedeafferenteerd gebied (Wall et al., 1986; Florence en Kaas, 1995). In het primaatmodel wordt de gedenerveerde cortex, 1 jaar na zenuwdoorsnijding en chirurgisch herstel, gekenmerkt door onvolledige en wanordelijke representaties van de geregenereerde en aangrenzende (intacte) zenuwen. Deze fragmentarische weergave wordt toegeschreven aan onvolledige perifere regeneratie, wat resulteert in een gedeeltelijk herstel van de gedenerveerde corticale ruimte (Kaas, 1991). Om de mate van perifere regeneratie in onze patiëntenpopulatie te beoordelen, hebben we sensorische en motorische zenuwgeleidingsonderzoeken uitgevoerd over het doorgesneden gebied. Onze zenuwgeleidingsresultaten tonen aan dat PNIr-patiënten een significant verminderde amplitude en verhoogde latentie in zowel sensorische als motorische zenuwen hebben in vergelijking met hun eigen niet-doorgesneden zijde. Verminderde amplitude gecombineerd met verhoogde latentie is indicatief voor verlies van perifere vezels (dwz celdood) en/of abnormale of onvolledige hermyelinisatie na transsectie (Kimura, 1984). Bovendien is het goed vastgesteld dat tussen 20% en 50% van de ganglionneuronen van de dorsale wortel afsterven na zenuwdoorsnijding (Liss et al., 1996). Afferente celdood en onvolledige regeneratie kunnen dus resulteren in verminderde afferente input naar de cortex, wat verantwoordelijk kan zijn voor aanhoudende sensorische tekorten en verminderde BOLD-respons in BA2 en S2. Bovendien zou deze verminderde afferente input ook de corticale verdunning kunnen verklaren die we in dezelfde regio's van de cortex hebben waargenomen. Van sensorische deprivatie is aangetoond dat het transneuronale degeneratie veroorzaakt in verschillende regio's van het CZS, inclusief de dorsale hoorn na een heupzenuwsectie (Knyihar-Csillik et al., 1989), en kan neuronen van de tweede en derde orde omvatten (Powell en Erulkar, 1962). Transneuronale degeneratie wordt gekenmerkt door celkrimp en er wordt gedacht dat het verband houdt met verminderde of niet-bestaande afferente input (Knyihar-Csillik et al., 1989). Corticaal verlies van grijze stof (of atrofie) zou dus ook direct gerelateerd kunnen zijn aan verminderde afferente input.
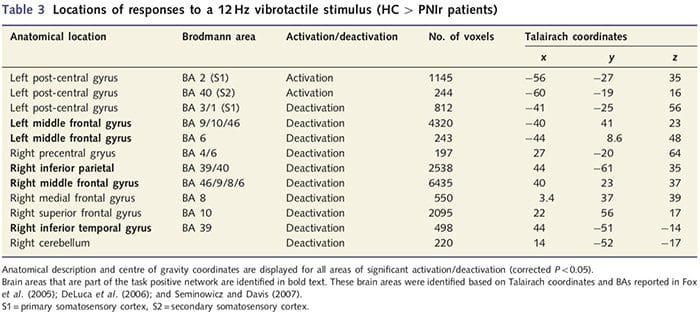
 We hebben ook verhoogde activering aangetoond in de post-centrale gyrus in een regio die overeenkomt met BA1/3 (Talairach en Tournoux, 1988). Elektrofysiologische, anatomische tracerings- en neuroimaging-onderzoeken hebben aangetoond dat voor de meeste cutane mechanoreceptieve afferenten de eerste corticale bestemmingen BA1 en BA3b zijn. Deze cytoarchitectonische hersengebieden hebben elk een somatotopische lichaamskaart met kleine receptieve velden. Bovendien reageren deze gebieden op veel kenmerken van tactiele informatie, zoals textuur en ruwheid, snelheid en kromming van stimuli (Bodegard et al., 2001). fMRI-onderzoeken hebben aangetoond dat activiteit in de somatosensorische cortex wordt beïnvloed door aandacht, zodat fMRI-reacties op tactiele stimuli in S1 worden verhoogd wanneer proefpersonen aandacht besteden aan een tactiele stimulus, maar worden verzwakt wanneer proefpersonen worden afgeleid (Arthurs et al., 2004; Porro et al., 2004). Bovendien activeerden onze patiënten een netwerk van hersengebieden dat bekend staat als het taakpositieve netwerk (DeLuca et al., 2006) meer dan gezonde controles. Deze hersengebieden worden geactiveerd tijdens processen die aandacht vragen (Fox et al., 2005; Seminowicz en Davis, 2007). Samen impliceren deze bevindingen dat patiënten vanwege hun verminderde sensorische input meer aandacht moeten besteden aan de stimulus dan aan de controlegroep. Deze verhoogde aandacht kan ook de verhoogde activering in BA1/3b verklaren. Natuurlijk kan de verhoogde activering in BA1/3b ook een weerspiegeling zijn van plasticiteit die geen verband houdt met aandachtsbelasting.
We hebben ook verhoogde activering aangetoond in de post-centrale gyrus in een regio die overeenkomt met BA1/3 (Talairach en Tournoux, 1988). Elektrofysiologische, anatomische tracerings- en neuroimaging-onderzoeken hebben aangetoond dat voor de meeste cutane mechanoreceptieve afferenten de eerste corticale bestemmingen BA1 en BA3b zijn. Deze cytoarchitectonische hersengebieden hebben elk een somatotopische lichaamskaart met kleine receptieve velden. Bovendien reageren deze gebieden op veel kenmerken van tactiele informatie, zoals textuur en ruwheid, snelheid en kromming van stimuli (Bodegard et al., 2001). fMRI-onderzoeken hebben aangetoond dat activiteit in de somatosensorische cortex wordt beïnvloed door aandacht, zodat fMRI-reacties op tactiele stimuli in S1 worden verhoogd wanneer proefpersonen aandacht besteden aan een tactiele stimulus, maar worden verzwakt wanneer proefpersonen worden afgeleid (Arthurs et al., 2004; Porro et al., 2004). Bovendien activeerden onze patiënten een netwerk van hersengebieden dat bekend staat als het taakpositieve netwerk (DeLuca et al., 2006) meer dan gezonde controles. Deze hersengebieden worden geactiveerd tijdens processen die aandacht vragen (Fox et al., 2005; Seminowicz en Davis, 2007). Samen impliceren deze bevindingen dat patiënten vanwege hun verminderde sensorische input meer aandacht moeten besteden aan de stimulus dan aan de controlegroep. Deze verhoogde aandacht kan ook de verhoogde activering in BA1/3b verklaren. Natuurlijk kan de verhoogde activering in BA1/3b ook een weerspiegeling zijn van plasticiteit die geen verband houdt met aandachtsbelasting.
 BA2 en S2 ontvangen beide projecties van BA1/3b en ook van verschillende delen van het ventroposterieure thalamische complex (Pons et al., 1985; Friedman en Murray, 1986). Allebei hersenen gebieden hebben grote, vaak meercijferige (BA2) of bilaterale (S2) receptieve velden (Pons et al., 1985; Iwamura et al., 2002). Op basis van anatomische projecties en neuronale responseigenschappen is hiërarchische verwerking van tactiele informatie aangetoond van BA1/3b tot BA 2 (Kaas et al., 2002). Bovendien suggereren elektrofysiologische studies bij makaken (Pons et al., 1987) en magneto-encefalografiegegevens verkregen bij mensen, dat seriële verwerking van tactiele inputs plaatsvindt van S1 tot S2 bij hogere primaten (Frot en Mauguiere 1999; Disbrow et al. ., 2001). Verschillende onderzoeken hebben aangetoond dat BA2 bij voorkeur wordt geactiveerd door vorm en kromming (Bodegard et al., 2001), terwijl S2 mogelijk betrokken is bij tactiel leren (Ridley en Ettlinger 1976; Murray en Mishkin, 1984), wat het idee ondersteunt dat deze hersengebieden zijn betrokken bij somatosensorische verwerking van hogere orde. Onze psychofysische beoordeling toonde aan dat patiënten significant verslechterd waren bij de detectie van eenvoudige tactiele stimuli, en in de Shape Texture Identification-test 1.5 jaar na de operatie (Taylor et al., 2008a). Deze laatste test beoordeelt het vermogen van een patiënt om kenmerken van een object te herkennen terwijl hij actief een vorm of textuur verkent, waarvoor de integratie van sensorische informatie over delen van het lichaam nodig is (Rosen en Lundborg, 1998). Alles bij elkaar genomen is een interpretatie van onze gegevens dat PNIr-patiënten meer aandacht besteden aan de vibrotactiele stimulus, wat leidt tot een verhoogde activering van het taak-positieve netwerk en BA1/3. Bij deze patiënten impliceren onze gegevens echter dat verwerkingsgebieden van hogere orde, zoals BA2 en S2, geen tactiele informatie hebben ontvangen, wat op zijn beurt kan resulteren in corticale verdunning en verminderde BOLD-reacties.
BA2 en S2 ontvangen beide projecties van BA1/3b en ook van verschillende delen van het ventroposterieure thalamische complex (Pons et al., 1985; Friedman en Murray, 1986). Allebei hersenen gebieden hebben grote, vaak meercijferige (BA2) of bilaterale (S2) receptieve velden (Pons et al., 1985; Iwamura et al., 2002). Op basis van anatomische projecties en neuronale responseigenschappen is hiërarchische verwerking van tactiele informatie aangetoond van BA1/3b tot BA 2 (Kaas et al., 2002). Bovendien suggereren elektrofysiologische studies bij makaken (Pons et al., 1987) en magneto-encefalografiegegevens verkregen bij mensen, dat seriële verwerking van tactiele inputs plaatsvindt van S1 tot S2 bij hogere primaten (Frot en Mauguiere 1999; Disbrow et al. ., 2001). Verschillende onderzoeken hebben aangetoond dat BA2 bij voorkeur wordt geactiveerd door vorm en kromming (Bodegard et al., 2001), terwijl S2 mogelijk betrokken is bij tactiel leren (Ridley en Ettlinger 1976; Murray en Mishkin, 1984), wat het idee ondersteunt dat deze hersengebieden zijn betrokken bij somatosensorische verwerking van hogere orde. Onze psychofysische beoordeling toonde aan dat patiënten significant verslechterd waren bij de detectie van eenvoudige tactiele stimuli, en in de Shape Texture Identification-test 1.5 jaar na de operatie (Taylor et al., 2008a). Deze laatste test beoordeelt het vermogen van een patiënt om kenmerken van een object te herkennen terwijl hij actief een vorm of textuur verkent, waarvoor de integratie van sensorische informatie over delen van het lichaam nodig is (Rosen en Lundborg, 1998). Alles bij elkaar genomen is een interpretatie van onze gegevens dat PNIr-patiënten meer aandacht besteden aan de vibrotactiele stimulus, wat leidt tot een verhoogde activering van het taak-positieve netwerk en BA1/3. Bij deze patiënten impliceren onze gegevens echter dat verwerkingsgebieden van hogere orde, zoals BA2 en S2, geen tactiele informatie hebben ontvangen, wat op zijn beurt kan resulteren in corticale verdunning en verminderde BOLD-reacties.
Men denkt dat de insula een rol speelt bij het integreren van multimodale informatie die belangrijk is voor sensomotorische, emotionele, allostatische/homeostatische en cognitieve functies (Devinsky et al., 1995; Critchley, 2004; Craig, 2008) en is aangeduid als een limbische sensorische cortex (Craig , 2008). Verschillende onderzoeken hebben eilandactivering gerapporteerd als reactie op tactiele stimulatie (Gelnar et al., 1998;
Downar et al., 2002) en anatomische traceringsstudies bij primaten hebben aangetoond dat de insula wederzijds verbonden is met frontale, pariëtale en temporale lobben (Augustine, 1996). Bij onze patiënten was de rechter anterieure insula het enige corticale gebied dat significante corticale verdunning vertoonde in combinatie met verminderde fractionele anisotropiewaarden in de aangrenzende witte stof, wat suggereert dat de corticale verdunning in dit gebied geassocieerd is met een verlies van vezels die uitsteken naar of van deze structuur. De rechter anterieure insula is betrokken bij interoceptie omdat deze is gesitueerd om homeostatische input van het lichaam te integreren met motiverende, emotionele en sociale omstandigheden (Craig, 2008). Verder hebben Critchley et al. (2004) rapporteerden een correlatie tussen interoceptieve vaardigheden en het volume van de grijze stof van de rechter voorste insula. Gezien onze bevinding dat de patiënten grijze stof in de rechter voorste insula hebben verminderd, zou het interessant zijn om de interoceptieve mogelijkheden na perifere zenuwbeschadiging in een toekomstig onderzoek te beoordelen.
Alles bij elkaar genomen hebben we voor het eerst aangetoond dat functionele en structurele veranderingen aanwezig zijn in de menselijke hersenschors, 1.5 jaar na een volledige doorsnijding van een perifere zenuw van de bovenste ledematen die microchirurgisch werd gerepareerd. Bovendien duiden zenuwgeleidingsmetingen op onvolledige perifere regeneratie bij deze patiënten. Verder laten we zien dat corticale dikte gerelateerd is aan psychofysische herstelmaatregelen, in die zin dat dunnere cortex in BA2 en S2 geassocieerd was met een slechtere somatosensorische functie. Deze gegevens suggereren dat het herstel van normale functionele activeringskaarten direct geassocieerd is met de succesvolle regeneratie van perifere afferenten.
Keri S. Taylor,1,2 Dimitri J. Anastakis2,3,4 en Karen D. Davis1,2,3
1 Afdeling Hersenen, Beeldvorming en Gedrag - Systems Neuroscience, Toronto Western Research Institute, University Health Network, Toronto, Canada M5T258
2 Instituut voor Medische Wetenschappen, Universiteit van Toronto, Canada
3 Afdeling Chirurgie, Universiteit van Toronto, Canada
4 Resource Centre voor klinische studies, Toronto Western Research Institute, University Health Network, Toronto, Canada M5T2S8
Correspondentie met: Karen D. Davis, Ph.D.,
Afdeling Hersenen, Beeldvorming en Gedrag - Systems Neuroscience, Toronto Western Research Institute,
Toronto West-ziekenhuis,
Universitair gezondheidsnetwerk,
Kamer MP14-306, 399 Bathurst Street,
Toronto, Ontario,
Canada M5T 2S8
E-mail: kdavis@uhnres.utoronto.ca
Danksagung
De auteurs danken de heer Geoff Pope, Dr. Adrian Crawley, de heer Eugene Hlasny en de heer Keith Ta voor deskundige technische bijstand. De auteurs willen Dr. Peter Ashby en Mr. Freddy Paiz van de Toronto Western Hospital EMG Clinic bedanken voor het uitvoeren van de zenuwgeleidingstests en voor het geven van een deskundige beoordeling van de bevindingen. De auteurs bedanken ook Drs Dvali, Binhammer, Fialkov en Antonyshyn voor hun samenwerking met dit project. Dr. Davis is een Canada Research Chair in Brain and Behaviour (CIHR MOP 53304).
Financiering
The Physicians� Services Incorporated en een gezamenlijke startsubsidie van het University of Toronto Centre for the Study of Pain/AstraZeneca.
Aanvullend materiaal
Aanvullend materiaal is beschikbaar op Brain online.
blanco
Referenties:
Apkarian AV, Sosa Y, Sonty S, et al. Chronische rugpijn wordt geassocieerd met
verminderde prefrontale en thalamische grijze stofdichtheid. J Neurosci
2004; 24: 10410.
Arthurs PB, Johansen-Berg H, Matthews PM, Boniface SJ. Aandacht
differentieel moduleert de koppeling van fMRI BOLD en evoked
potentiële signaalamplitudes in de menselijke somatosensorische cortex 7.
Exp Hersenonderzoek 2004; 157: 269.
Augustinus Jr. Circuits en functionele aspecten van de insulaire kwab
bij primaten, inclusief mensen. Hersenonderzoek Hersenonderzoek Rev 1996; 22:
229�44.
Beaulieu C. De basis van anisotrope waterdiffusie in het zenuwstelsel
systeem – een technische beoordeling. NMR Biomed 2002; 15: 435-55.
Bodegard A, Geyer S, Grefkes C, Zilles K, Roland PE. hiërarchisch
verwerking van tactiele vorm in het menselijk brein. Neuron 2001; 31:
317�28.
Craig AD. Interoceptie en emotie: een neuroanatomisch perspectief.
In: Lewis M, Haviland-Jones J, Barrett L, redacteuren. Handboek van
emoties. New York: Guildford Press; 2008. blz. 272.
Critchley HD. De menselijke cortex reageert op een interoceptieve uitdaging.
Proc Natl Acad Sci VS 2004; 101: 6333.
Critchley HD, Wiens S, Rotshtein P, Ohman A, Dolan RJ. neurale systemen
het ondersteunen van interoceptief bewustzijn. Nat Neurosci 2004; 7: 189.
Dale AM, Fischl B, Sereno MI. Corticale oppervlakte-gebaseerde analyse. I. Segmentatie
en oppervlaktereconstructie. Neurobeeld 1999; 9: 179.
Davis KD, Pope G, Chen J, Kwan CL, Crawley AP, Diamant NE. Corticaal
dunner worden bij IBS: implicaties voor homeostase, aandacht en pijn
verwerken. Neurologie 2008; 70: 153.
DeLuca M, Beckmann CF, De SN, Matthews PM, Smith SM. fMRI in rust
staatsnetwerken definiëren verschillende manieren van interactie over lange afstanden
in het menselijk brein. Neurobeeld 2006; 29: 1359-67.
Devinsky O, Morrell MJ, Vogt BA. Bijdragen van anterieure cingulate
cortex naar gedrag. Hersenen 1995; 118 (Pt 1): 279.
Disbrow E, Roberts T, Poeppel D, Krubitzer L. Bewijs voor interhemisferisch
verwerking van invoer van de handen in de mens S2
en PV. J Neurofysiol 2001; 85: 2236�44.
Downar J, Crawley AP, Mikulis DJ, Davis KD. Een corticaal netwerkgevoelig
om opvallendheid te stimuleren in een neutrale gedragscontext over meerdere
zintuiglijke modaliteiten. J Neurofysiologie 2002; 87: 615-20.
Draganski B, Moser T, Lummel N, et al. Afname van thalamisch grijs
kwestie na amputatie van een ledemaat. Neurobeeld 2006; 31: 951�7.
Fischl B, Dale AM. De dikte van de menselijke hersenschors meten
van magnetische resonantiebeelden. Proc Natl Acad Sci VS 2000; 97:
11050�5.
Fischl B, Sereno MI, Dale AM. Corticale oppervlakte-gebaseerde analyse. II:
Inflatie, afvlakking en een op het oppervlak gebaseerd coördinatensysteem.
Neurobeeld 1999a; 9: 195-207.
Fischl B, Sereno MI, Tootell RB, Dale AM. Intersubject met hoge resolutie
middeling en een coördinatensysteem voor het corticale oppervlak. Hum Brein
Kaart 1999b; 8: 272.
Florence SL, Kaas JH. Grootschalige reorganisatie op meerdere niveaus van de
somatosensorische route volgt therapeutische amputatie van de hand
bij apen. J Neurosci 1995; 15: 8083.
Fox MD, Snyder AZ, Vincent JL, Corbetta M, Van E, Raichle ME. De
menselijk brein is intrinsiek georganiseerd in dynamische, antigecorreleerde
functionele netwerken. Proc Natl Acad Sci VS 2005; 102: 9673-9678.
Friedman DP, Murray EA. Thalamische connectiviteit van de tweede
somatosensorische gebied en aangrenzende somatosensorische velden van de
laterale sulcus van de makaak. J Comp Neurol 1986; 252: 348.
Frot M, Mauguiere F. Timing en ruimtelijke verdeling van somatosensorische
reacties geregistreerd in de bovenste oever van de Sylvian Fissure (SII-gebied) in
mensen. Cereb Cortex 1999; 9: 854�63.
Geha PY, Baliki MN, Harden RN, Bauer WR, Parrish TB, Apkarian AV.
De hersenen bij chronische CRPS-pijn: abnormale grijs-witte stof
interacties in emotionele en autonome regio's. Neuron 2008; 60:
570�81.
Gelnar PA, Krauss BR, Szeverenyi NM, Apkarian AV. Vingertopweergave
in de menselijke somatosensorische cortex: een fMRI-onderzoek.
Neurobeeld 1998; 7: 261.
Hickmott PW, Steen PA. Grootschalige veranderingen in dendritische structuur
tijdens reorganisatie van de volwassen somatosensorische cortex. Nat Neurosci
2005; 8: 140.
Iwamura Y, Tanaka M, Iriki A, Taoka M, Toda T. Verwerking van
tactiele en kinesthetische signalen van bilaterale zijden van het lichaam in de
postcentrale gyrus van wakkere apen. Gedrag Hersenonderzoek 2002; 135:
185�90.
Jaquet JB, Luijsterburg AJ, Kalmijn S, Kuypers PD, Hofman A, Hovius SE.
Mediane, ulnaire en gecombineerde mediaan-ulnaire zenuwletsels: functioneel
resultaat en terugkeer naar productiviteit. J Trauma 2001; 51: 687.
Jurkiewicz MT, Crawley AP, Verrier MC, Fehlings MG, Mikulis DJ.
Somatosensorische corticale atrofie na dwarslaesie: een voxelbased
morfometrie studie. Neurologie 2006; 66: 762�4.
Kaas JH. Plasticiteit van sensorische en motorische kaarten bij volwassen zoogdieren. annu
Rev Neurosci 1991; 14: 137.
Kaas JH, Jain N, Qi HX. De organisatie van het somatosensorische systeem in
primaten. In: Nelson RJ, redacteur. Het somatosensorische systeem.
Washington, DC: CRC Press; 2002. blz. 1�25.
Kimura J. Elektrodiagnose bij aandoeningen van zenuw en spier: principes
en oefenen. Oxford: Oxford University Press; 2001.
Kimura J. Principes en valkuilen van zenuwgeleidingsonderzoeken. Ann Neurol
1984; 16: 415.
Knyihar-Csillik E, Rakic P, Csillik B. Transneuronale degeneratie in
de Rolando-substantie van het ruggenmerg van primaten opgeroepen door
door axotomie geïnduceerde transganglionische degeneratieve atrofie van centrale
primaire sensorische terminals. Celweefselonderzoek 1989; 258: 515.
Liss AG, af Ekenstam FW, Wiberg M. Verlies van neuronen in de dorsale wortel
ganglia na doorsnijding van een perifere sensorische zenuw. een anatomische
onderzoek bij apen. Scand J Plast Reconstr Surg Hand Surg 1996; 30: 1�6.
Lotze M, Flor H, Grodd W, Larbig W, Birbaumer N. Phantom-bewegingen
en pijn. Een fMRI-onderzoek bij geamputeerden van de bovenste ledematen. Hersenen 2001;
124: 2268-77.
Manduch M, Bezuhly M, Anastakis DJ, Crawley AP, Mikulis DJ. serieel
fMRI van adaptieve veranderingen in de primaire sensomotorische cortex volgend
duim reconstructie. Neurologie 2002; 59: 1278.
May A. Chronische pijn kan de structuur van de hersenen veranderen. Pijn 2008;
137: 7-15.
McAllister RM, Gilbert SE, Calder JS, Smith PJ. De epidemiologie en
behandeling van perifere zenuwletsels van de bovenste ledematen in moderne
oefening. J Hand Surg (Br) 1996; 21: 4�13.
Murray EA, Mishkin M. Relatieve bijdragen van SII en area5 aan tactiele
discriminatie bij apen 2. Behav Brain Res 1984; 11: 67.
Napadow V, Kettner N, Ryan A, Kwong KK, Audette J, Hui KK.
Somatosensorische corticale plasticiteit bij carpaletunnelsyndroom - een dwarsdoorsnede
fMRI-evaluatie. Neurobeeld 2006; 31: 520�30.
Oldfield RC. De beoordeling en analyse van handigheid: de Edinburgh
voorraad. Neuropsychologie 1971; 9: 97�113.
Pons TP, Garraghty PE, Cusick CG, Kaas JH. De somatotopische organisatie
van gebied 2 bij makaken 6. J Comp Neurol 1985; 241: 445.
Pons TP, Garraghty PE, Friedman DP, Mishkin M. Fysiologisch bewijs
voor seriële verwerking in somatosensorische cortex. Wetenschap 1987; 237:
417�20.
Porro CA, Lui F, Facchin P, Maieron M, Baraldi P. Percept-gerelateerde activiteit
in het menselijke somatosensorische systeem: functionele magnetische resonantie
beeldvormende onderzoeken. Magn Reson Imaging 2004; 22: 1539-48.
Powell TP, Erulkar S. Transneuronale celdegeneratie in de auditieve
relaiskernen van de kat. J Anat 1962; 96: 249.
Ridley RM, Ettlinger G. Verminderd tactiel leren en retentie na
verwijderingen van de tweede somatische sensorische projectiecortex (SII) in de
aap. Hersenonderzoek 1976; 109: 656.
Rosen B, Lundborg G. Een nieuw instrument voor tactiele gnosis in gevoeligheid
testen. J Hand Ther 1998; 11: 251�7.
Seminowicz DA, Davis KD. Pijn verbetert functionele connectiviteit
van een hersennetwerk opgeroepen door het uitvoeren van een cognitieve taak.
J Neurofysiol 2007; 97: 3651.
Smith SM, Jenkinson M, Johansen-Berg H, et al. Op traktaat gebaseerd ruimtelijk
statistieken: voxelgewijze analyse van diffusiegegevens over meerdere onderwerpen.
Neurobeeld 2006; 31: 1487.
Talairach J, Tournoux P. Co-planaire stereotaxische atlas van het menselijk brein.
New York: Thieme Medical Publishers Inc.; 1988.
Taylor KS, Anastakis DJ, Davis KD. Chronische pijn na perifere
zenuwbeschadiging wordt geassocieerd met catastrofale pijn en neuroticisme.
Int Ass Stud Pain 2008a; 267.
Taylor KS, Davis KD. Stabiliteit van tactiele en pijngerelateerde fMRI
hersenactivaties: een onderzoek naar drempelafhankelijke en
drempelonafhankelijke methoden. Hum Brain Map 2009; 30:
1947�62.
Taylor KS, Seminowicz DA, Davis KD. Twee systemen van rusttoestand
verbinding tussen de insula en de cingulate cortex. Hum Brein
Kaart 2008b;DOI:10.1002/hbm.20705.
Wall JT, Kaas JH, Sur M, Nelson RJ, Felleman DJ, Merzenich MM.
Functionele reorganisatie in somatosensorische corticale gebieden 3b en 1
van volwassen apen na herstel van de mediane zenuw: mogelijk verband met
zintuiglijk herstel bij de mens. J Neurosci 1986; 6: 218.
Accordeon sluiten
Professionele reikwijdte van de praktijk *
De informatie hierin over "Je zenuw snijden verandert je hersenen | El Paso, TX." is niet bedoeld ter vervanging van een een-op-een relatie met een gekwalificeerde zorgverlener of gediplomeerde arts en is geen medisch advies. We moedigen u aan om beslissingen over de gezondheidszorg te nemen op basis van uw onderzoek en samenwerking met een gekwalificeerde zorgverlener.
Blog Informatie & Scope Discussies
Ons informatiebereik: is beperkt tot chiropractie, musculoskeletale, fysieke medicijnen, welzijn, bijdragende etiologische viscerosomatische stoornissen binnen klinische presentaties, geassocieerde somatoviscerale reflex klinische dynamiek, subluxatiecomplexen, gevoelige gezondheidskwesties en/of functionele geneeskunde artikelen, onderwerpen en discussies.
Wij bieden en presenteren klinische samenwerking met specialisten uit verschillende disciplines. Elke specialist wordt beheerst door hun professionele praktijk en hun jurisdictie van licentiestatus. We gebruiken functionele gezondheids- en welzijnsprotocollen om de verwondingen of aandoeningen van het bewegingsapparaat te behandelen en te ondersteunen.
Onze video's, berichten, onderwerpen, onderwerpen en inzichten behandelen klinische zaken, problemen en onderwerpen die verband houden met en direct of indirect onze klinische praktijk ondersteunen.*
Ons kantoor heeft redelijkerwijs geprobeerd om ondersteunende citaten te geven en heeft de relevante onderzoeksstudie of studies geïdentificeerd die onze berichten ondersteunen. Wij verstrekken kopieën van ondersteunende onderzoeksstudies die op verzoek beschikbaar zijn voor regelgevende instanties en het publiek.
We begrijpen dat we zaken behandelen die een aanvullende uitleg vereisen over hoe het kan helpen bij een bepaald zorgplan of behandelprotocol; daarom, om het onderwerp hierboven verder te bespreken, aarzel dan niet om te vragen Dr. Alex Jimenez, DC, of neem contact met ons op 915-850-0900.
Wij zijn er om u en uw gezin te helpen.
zegeningen
Dr. Alex Jimenez DC MSACP, RN*, CCST, IFMCP*, CIFM*, ATN*
e-mail: coach@elpasofunctionalmedicine.com
Licentie als Doctor of Chiropractic (DC) in Texas & New Mexico*
Texas DC-licentie # TX5807, New Mexico DC-licentie # NM-DC2182
Licentie als geregistreerde verpleegkundige (RN*) in Florida
Florida-licentie RN-licentie # RN9617241 (controle nr. 3558029)
Compacte status: Licentie voor meerdere staten: Geautoriseerd om te oefenen in 40 Staten*
Dr. Alex Jimenez DC, MSACP, RN* CIFM*, IFMCP*, ATN*, CCST
Mijn digitale visitekaartje





