
We hebben als kinderen allemaal geleerd dat er 5 zintuigen zijn: zien, proeven, klinken, ruiken en voelen. De eerste vier zintuigen maken gebruik van duidelijke, onderscheiden organen, zoals de ogen, smaakpapillen, oren en neus, maar hoe raakt het lichaamsgevoel precies aan? Aanraking wordt ervaren over het hele lichaam, zowel binnen als buiten. Er is niet één afzonderlijk orgaan dat verantwoordelijk is voor het voelen van aanraking. In plaats daarvan zijn er kleine receptoren, of zenuwuiteinden, rond het hele lichaam die aanraking voelen waar het plaatsvindt en signalen naar de hersenen sturen met informatie over het type aanraking dat heeft plaatsgevonden. Zoals een smaakpapillen op de tong smaak detecteert, zijn mechanoreceptoren klieren in de huid en op andere organen die sensaties van aanraking detecteren. Ze staan bekend als mechanorecep- omdat ze zijn ontworpen om mechanische sensaties of drukverschillen te detecteren.
Inhoud
Rol van Mechanoreceptoren
Een persoon begrijpt dat hij een sensatie heeft ervaren zodra het orgaan dat verantwoordelijk is voor het ontdekken van dat specifieke zintuig een bericht naar de hersenen stuurt, het primaire orgaan dat alle informatie verwerkt en ordent. Berichten worden vanuit alle delen van het lichaam naar de hersenen gestuurd via draden die neuronen worden genoemd. Er zijn duizenden kleine neuronen die zich vertakken naar alle delen van het menselijk lichaam, en aan de uiteinden van veel van deze neuronen bevinden zich mechanoreceptoren. Om te laten zien wat er gebeurt als je een object aanraakt, gebruiken we een voorbeeld.
Stel je voor dat er een mug op je arm landt. De stam van dit insect, zo licht, stimuleert mechanoreceptoren in dat specifieke deel van de arm. Die mechanoreceptoren sturen een bericht langs het neuron waarmee ze zijn verbonden. Het neuron maakt helemaal verbinding met de hersenen, die het bericht ontvangen dat iets je lichaam aanraakt op de exacte locatie van de specifieke mechanoreceptor die het bericht heeft verzonden. De hersenen zullen handelen met dit advies. Misschien zal het de ogen vertellen om naar het deel van de arm te kijken dat de handtekening heeft gedetecteerd. En wanneer de ogen de hersenen vertellen dat er een mug op de arm zit, kunnen de hersenen de hand vertellen om hem snel weg te vegen. Dat is hoe mechanoreceptoren werken. Het doel van het onderstaande artikel is om de functionele organisatie en moleculaire determinanten van mechanoreceptoren aan te tonen en in detail te bespreken.
Touch Sense: functionele organisatie en moleculaire determinanten van mechanisch gevoelige receptoren
Abstract
Mechanoreceptoren van de huid zijn gelokaliseerd in de verschillende lagen van de huid waar ze een breed scala aan mechanische stimuli detecteren, waaronder lichte borstel, rek, trillingen en schadelijke druk. Deze verscheidenheid aan stimuli wordt geëvenaard door een diverse reeks gespecialiseerde mechanoreceptoren die op een specifieke manier reageren op huidvervorming en deze stimuli doorgeven aan hogere hersenstructuren. Studies over mechanoreceptoren en genetisch traceerbare sensorische zenuwuiteinden beginnen mechanismen voor aanrakingsgevoeligheid bloot te leggen. Werk op dit gebied heeft onderzoekers een beter begrip gegeven van de circuitorganisatie die ten grondslag ligt aan de perceptie van aanraking. Nieuwe ionenkanalen zijn naar voren gekomen als kandidaten voor transductiemoleculen en eigenschappen van mechanisch gestuurde stromen verbeterden ons begrip van de mechanismen van aanpassing aan tactiele stimuli. Deze recensie benadrukt de vooruitgang die is geboekt bij het karakteriseren van functionele eigenschappen van mechanoreceptoren in harige en kale huid en ionenkanalen die mechanische inputs detecteren en mechanoreceptoraanpassing vormgeven.
sleutelwoorden: mechanoreceptor, mechanosensitieve kanaal, pijn, huid, somatosensorisch systeem, aanraking
Introductie
Aanraking is de detectie van mechanische prikkels die de huid beïnvloeden, inclusief onschadelijke en schadelijke mechanische prikkels. Het is een essentieel zintuig voor het voortbestaan en de ontwikkeling van zoogdieren en mensen. Contact van vaste voorwerpen en vloeistoffen met de huid geeft de nodige informatie aan het centrale zenuwstelsel die verkenning en herkenning van de omgeving mogelijk maakt en voortbeweging of geplande handbeweging initieert. Aanraking is ook erg belangrijk voor leertijd, sociale contacten en seksualiteit. De tastzin is het minst kwetsbare zintuig, hoewel het bij veel pathologische aandoeningen kan worden vervormd (hyperesthesie, hypesthesie).1-3
Aanraakreacties omvatten een zeer nauwkeurige codering van mechanische informatie. Mechanoreceptoren van de huid zijn gelokaliseerd in de verschillende lagen van de huid waar ze een breed scala aan mechanische stimuli detecteren, waaronder lichte borstel, rek, trillingen, afbuiging van het haar en schadelijke druk. Deze verscheidenheid aan stimuli wordt geëvenaard door een diverse reeks gespecialiseerde mechanoreceptoren die op een specifieke manier reageren op huidvervorming en deze stimuli doorgeven aan hogere hersenstructuren. Somatosensorische neuronen van de huid vallen in twee groepen uiteen: laagdrempelige mechanoreceptoren (LTMR's) die reageren op goedaardige druk en hoogdrempelige mechanoreceptoren (HTMR's) die reageren op schadelijke mechanische stimulatie. LTMR- en HTMR-cellichamen bevinden zich in dorsale wortelganglia (DRG) en craniale sensorische ganglia (trigeminale ganglia). Zenuwvezels geassocieerd met LTMR's en HTMR's worden geclassificeerd als A?-, A?- of C-vezels op basis van hun actiepotentiaalgeleidingssnelheden. C-vezels zijn niet-gemyeliniseerd en hebben de langzaamste geleidingssnelheden (~2 m/s), terwijl A? en een? vezels zijn licht en zwaar gemyeliniseerd en vertonen respectievelijk intermediaire (~12 m/s) en snelle (~20 m/s) geleidingssnelheden. LTMR's worden ook geclassificeerd als langzaam of snel aanpassende reacties (SA- en RA-LTMR's) op basis van hun mate van aanpassing aan aanhoudende mechanische stimulus. Ze onderscheiden zich verder door de cutane eindorganen die ze innerveren en hun voorkeursstimuli.
Het vermogen van mechanoreceptoren om mechanische signalen te detecteren, is afhankelijk van de aanwezigheid van mechanotransducer-ionkanalen die mechanische krachten snel omzetten in elektrische signalen en het receptieve veld depolariseren. Deze lokale depolarisatie, receptorpotentiaal genoemd, kan actiepotentialen genereren die zich naar het centrale zenuwstelsel voortplanten. De eigenschappen van moleculen die mechanotransductie en aanpassing aan mechanische krachten bemiddelen, blijven echter onduidelijk.
In deze review geven we een overzicht van de mechanoreceptor-eigenschappen van zoogdieren bij onschadelijke en schadelijke aanrakingen in de harige en kale huid. We beschouwen ook de recente kennis over de eigenschappen van mechanisch gestuurde stromen in een poging het mechanisme van de aanpassing van de mechanoreceptor te verklaren. Ten slotte bekijken we de recente vooruitgang die is geboekt bij het identificeren van ionenkanalen en bijbehorende eiwitten die verantwoordelijk zijn voor het genereren van mechano-gated stromen.
Onschadelijke aanraking
Haarfollikel-geassocieerde LTMR's
De haarzakjes vertegenwoordigen haarschacht-producerende mini-organen die lichte aanraking detecteren. Vezels geassocieerd met haarzakjes reageren op haarbeweging en de richting ervan door reeksen actiepotentialen af te vuren bij het begin en verwijdering van de stimulus. Ze passen zich snel aan receptoren aan.
Kat en konijn. In katten- en konijnenvacht kunnen haarzakjes worden onderverdeeld in drie haarzakjes, het donshaar, het wachthaar en de tylotrichs. De donsharen (onderhaar, wol, vellus)4 zijn de meest talrijke, de kortste en fijnste haren van de vacht. Ze zijn golvend, kleurloos en kwamen in groepen van twee tot vier haren uit een gemeenschappelijke opening in de huid. De wachtharen (monotrichs, afluisteren, tophaar)4 zijn licht gebogen, al dan niet gepigmenteerd, en kwamen afzonderlijk uit de monden van hun follikels. De tylotrichs zijn het minst talrijk, de langste en dikste haren.5,6 Ze zijn gepigmenteerd of ongepigmenteerd, soms beide en kwamen afzonderlijk voort uit een follikel die omgeven is door een lus van capillaire bloedvaten. De sensorische vezels die een haarzakje leveren, bevinden zich onder de talgklier en worden toegeschreven aan A? of A?-LTMR-vezels.7
Dicht bij de donshaarschacht, net onder het niveau van de talgklier, bevindt zich de ring van lancetvormige pilo-Ruffini-uiteinden. Deze sensorische zenuwuiteinden bevinden zich in een spiraalvormige baan rond de haarschacht in het bindweefsel dat de haarfollikel vormt. Binnen de haarfollikel zijn er ook vrije zenuwuiteinden, waarvan sommige mechanoreceptoren vormen. Vaak omringen bloedlichaampjes (zie kale huid) het nekgebied van de tylotrich follikel.
Eigenschappen van gemyeliniseerde zenuwuiteinden in de harige huid van katten en konijnen zijn intensief onderzocht in de periode 1930-1970 (overzicht in Hamann, 1995).8 Opmerkelijk is dat Brown en Iggo 772 eenheden bestudeerden met gemyeliniseerde afferente zenuwvezels in de sapheneuze zenuwen van en konijn, hebben reacties geclassificeerd in drie receptortypes die overeenkomen met de bewegingen van donshaar (type D-receptoren), bewakingshaar (type G-receptoren) en Tylotrich-haar (type T-receptor).9 Alle afferente zenuwvezelreacties zijn samengebracht in de snel aanpassende receptor van type I (RA I) door oppositie tegen de Pacinische receptor genaamd RA II. RA I-mechanoreceptoren detecteren de snelheid van mechanische stimulus en hebben een scherpe grens. Ze detecteren geen thermische variaties. Burgess et al. beschreef ook een snel aanpassende veldreceptor die optimaal reageert op het aaien van de huid of beweging van meerdere haren, wat werd toegeschreven aan stimulatie van pilo-Ruffini-uiteinden. Geen van de haarzakjesreacties werd toegeschreven aan C-vezelactiviteit
Muizen. In de dorsale harige huid van muizen zijn drie hoofdtypen haarzakjes beschreven: zigzag (ongeveer 72%), priem/auchene (ongeveer 23%) en bewaker of tylotrich (ongeveer 5%).11-14 Zigzag en priem/ auchenne haarzakjes produceren de dunnere en kortere haarschachten en zijn geassocieerd met één talgklier. Guard of tylotrich haren zijn de langste van de haarzakjes. Ze worden gekenmerkt door een grote haarbol geassocieerd met twee talgklieren. Guard en priem/auchene haren zijn gerangschikt in een iteratief, regelmatig uit elkaar geplaatst patroon, terwijl zigzag haren dichtbevolkte huidgebieden rond de twee grotere haarzakjes typen [Fig. 1 (A1, A2 en A3)].
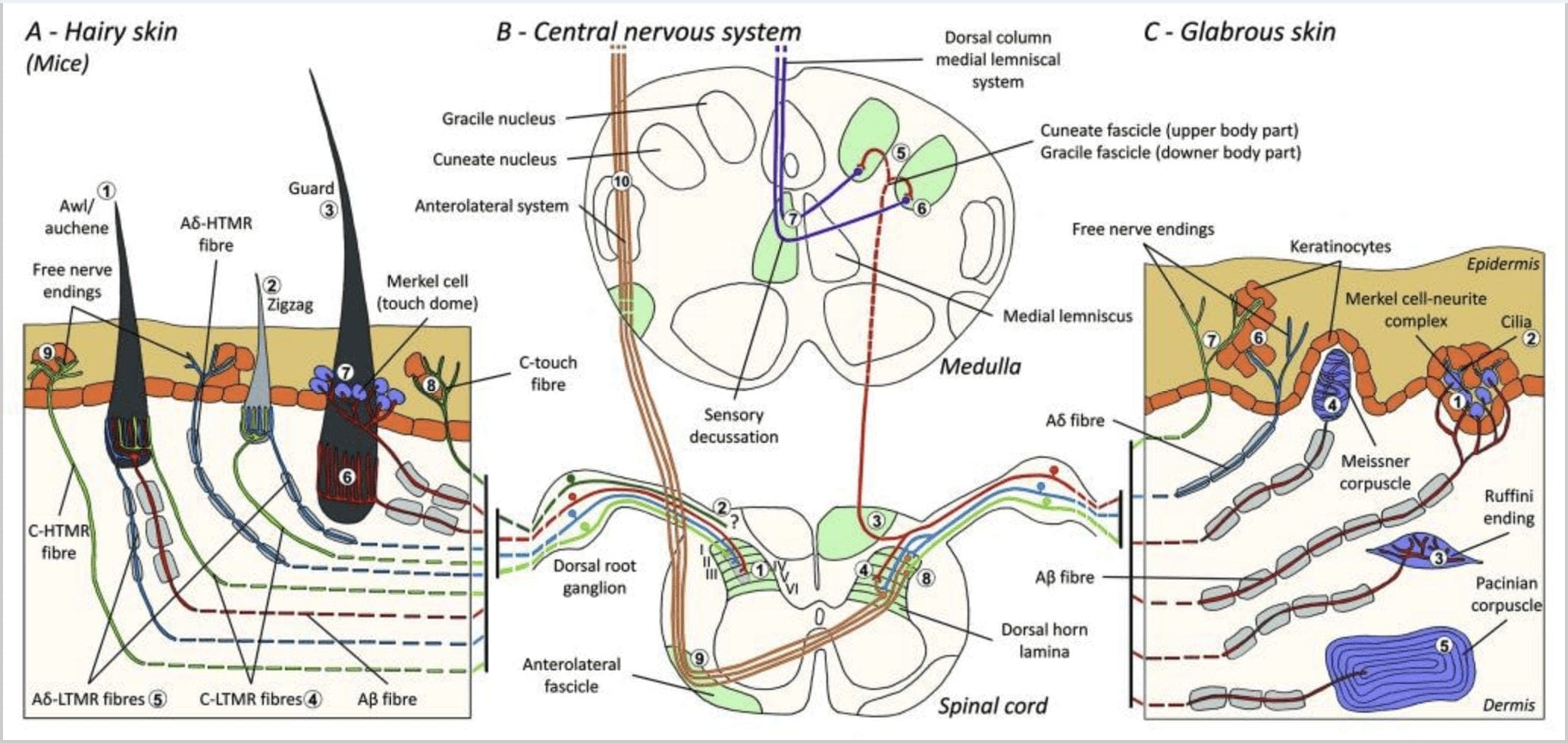
Figuur 1. Organisatie en projecties van cutane mechanoreceptoren. Bij een behaarde huid wordt licht borstelen en aanraken vooral gedetecteerd door de innervatie rond de haarzakjes: priem/auchenne (A1), zigzag (A2) en beschermer (A3). Priem/auchene haren worden driemaal geïnnerveerd door C-LTMR lancetvormige uiteinden (A4), A?-LTMR en A? snel aanpassend-LTMR (A6). Zigzag-haarzakjes zijn de kortere haarschachten en worden geïnnerveerd door zowel C-LTMR (A4) als A? -LTMR lancetvormige uiteinden (A5). De langste wachthaarzakjes worden geïnnerveerd door A? snel aanpassende LTMR longitudinale lancetvormige uiteinden (A6) en zijn geassocieerd met A? langzaam aanpassende LTMR van touch dome-uiteinden (A7). De centrale uitsteeksels van al deze vezels eindigen in afzonderlijke, maar gedeeltelijk overlappende laminae van de dorsale hoorn van het ruggenmerg (C-LTMR in lamina II, A?-LTMR in lamina III en A?-LTMR in lamina IV en V). De projecties van LTMR die dezelfde of aangrenzende haarzakjes innerveren, zijn uitgelijnd om een smalle kolom te vormen in de dorsale hoorn van het ruggenmerg (B1 in grijs). Alleen bij een harige huid innerveert een subpopulatie van C-vezels met vrije uiteinden de opperhuid en reageert op aangename aanraking (A8). Deze C-touch-vezels reageren niet op schadelijke aanraking en hun traject is nog niet bekend (B2). Bij een kale huid wordt onschadelijke aanraking gemedieerd door vier soorten LTMR's. Het Merkel-cel-neurietcomplex bevindt zich in de basale laag van de epidermis (C1). Deze mechanoreceptor bestaat uit een opstelling tussen veel Merkel-cellen en een vergrote zenuwuiteinde van een enkele A? vezel. Merkelcellen vertonen vingerachtige processen die in contact komen met keratinocyten (C2). Het Ruffini-uiteinde is gelokaliseerd in de dermis. Het is een dunne sigaarvormige ingekapselde zintuiglijke uiteinden verbonden met A? vezel (C3). Het bloedlichaampje van Meissner verbonden met A? zenuwuiteinde en bevindt zich in de dermale papillen. Deze ingekapselde mechanoreceptor bestaat uit opeengepakte ondersteunende cellen gerangschikt als horizontale lamellen omgeven door bindweefsel (C4). Pacinisch bloedlichaampje is de diepere mechanoreceptor. Een enkele A? niet-gemyeliniseerde zenuwuiteinde eindigt in het midden van dit grote eivormige lichaampje gemaakt van concentrische lamellen. Uitsteeksels van deze A?-LTMR-vezels in het ruggenmerg zijn verdeeld in twee takken. De belangrijkste centrale tak (B3) stijgt in het ruggenmerg in de ipsilaterale dorsale en vormt cuneate of gracile bundels (B5) op medulla-niveau waar de primaire afferenten hun eerste synaps maken (B6). De secundaire neuronen maken een sensorische decussatie (B7) om een kanaal te vormen op de mediale lemniscus die opstijgt door de hersenstam naar de middenhersenen, met name in de thalamus. Secundaire tak van LTMR eindigt in de dorsale hoorn in de lamina II, IV, V en interfereert met de pijnoverdracht (B4). Schadelijke aanraking wordt gedetecteerd door de vrije zenuw die eindigt in de epidermis van zowel de harige (A9) als de kale huid (C7). Deze mechanoreceptoren zijn het einde van A?-HTMR en C-HTMR in nauw contact met naburige keratinocyten (C6). AP-hTMR eindigen in de lamina I en V; C-HTMR eindigen in de lamina I en II (B8). Op het niveau van de dorsale hoorn van het ruggenmerg maken primaire afferente HTMR's synapsen met secundaire neuronen die de middellijn kruisen en naar de hogere hersenstructuur in de anterolaterale fascikel klimmen (B9, B10).
Onlangs hebben Ginty en medewerkers een combinatie van moleculair-genetische labeling en somatotopische retrograde traceringsbenaderingen gebruikt om de organisatie van perifere en centrale axonale uiteinden van de LTMR's bij muizen te visualiseren.15 Hun bevindingen ondersteunen een model waarin individuele kenmerken van een complexe tactiele stimulus worden geëxtraheerd door de drie haarfollikeltypen en via de activiteiten van unieke combinaties van A?-, A?- en C-vezels naar de dorsale hoorn getransporteerd.
Ze toonden aan dat de genetische labeling van tyrosinehydroxylase-positieve (TH+) DRG-neuronen een populatie van niet-peptiderge sensorische neuronen met een kleine diameter karakteriseren en visualisatie van C-LTMR perifere uiteinden in de huid mogelijk maken. Verrassend genoeg bleken de axonale takken van individuele C-LTMR's bomen te vormen en longitudinale lancetvormige uiteinden te vormen die nauw verbonden zijn met zigzag (80% van de uiteinden) en priem / auchene (20% van de uiteinden), maar geen tylotrich haarzakjes [Fig. 1 (A4)]. Lang werd gedacht dat longitudinale lancetvormige uiteinden uitsluitend tot A?-LTMR's behoorden en daarom was het onverwacht dat de uiteinden van C-LTMR's longitudinale lancetvormige uiteinden zouden vormen.15 Deze C-LTMR's hebben een intermediaire aanpassing in vergelijking met de langzaam en snel aanpassende gemyeliniseerde mechanoreceptoren [Fig. 2 (C1)].
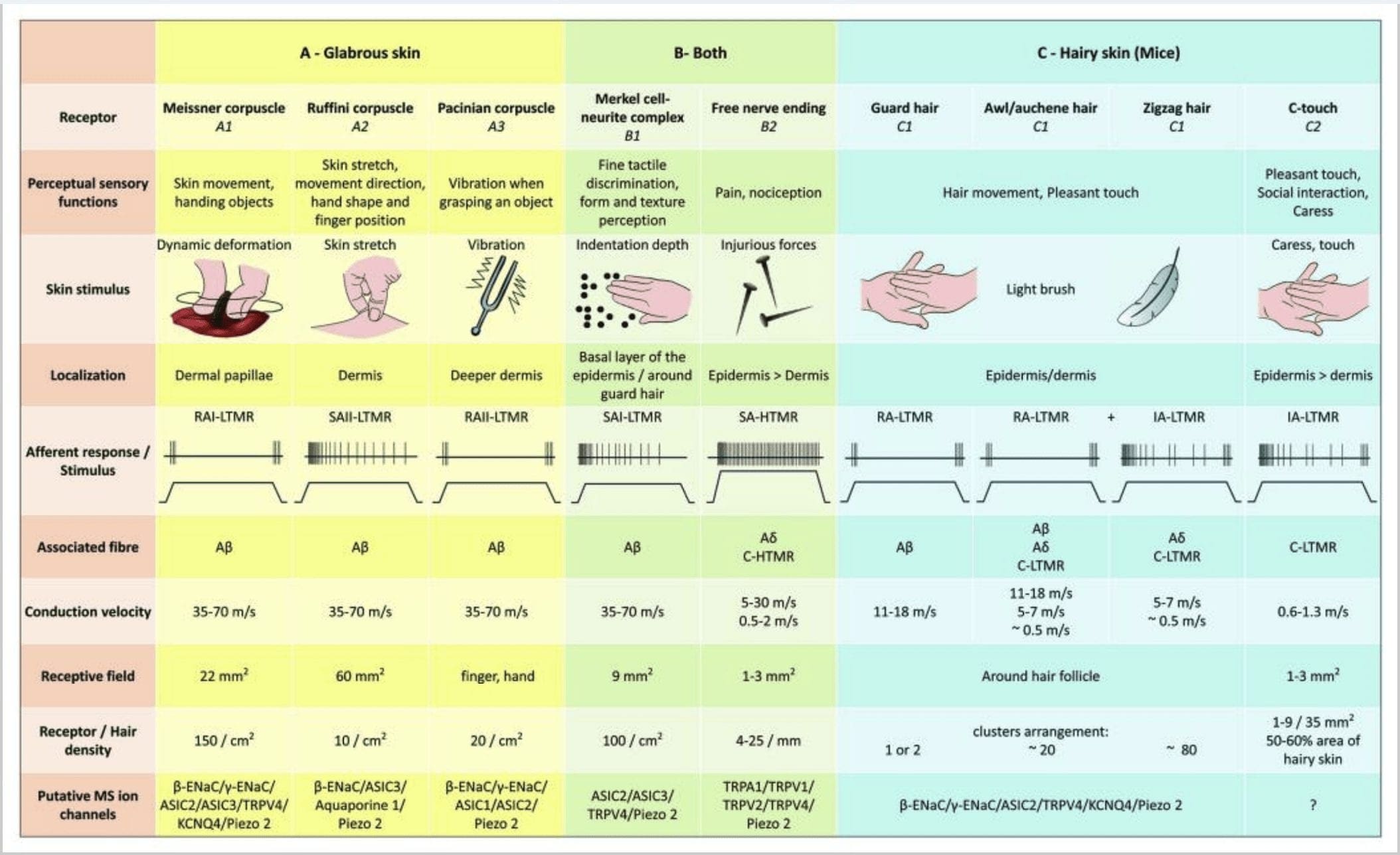
Figuur 2. Tactiele receptoren bij zoogdieren: Cutane tactiele receptoren differentiëren in onschadelijke aanraking ondersteund door meerdere receptoren met een lage mechanische drempel (LTMR's) in een kale en harige huid en schadelijke aanraking ondersteund door hoge mechanische drempelreceptor (HTMR's). Ze vormen zenuwvrije uiteinden die voornamelijk eindigen in de epidermis. (A) Gladde huid. A1: Bloedlichaampjes van Meissner detecteren huidbewegingen en wegglijden van voorwerpen in de hand. Ze zijn belangrijk voor het overhandigen van voorwerpen en behendigheid. Receptoren passen zich snel aan stimulus aan, zijn verbonden met A? vezels en dun tot C-vezels en hebben een groot receptorveld. A2: De bloedlichaampjes van Ruffini detecteren huidrek en zijn belangrijk om de vingerpositie en het overhandigen van een voorwerp te detecteren. Receptor past zich langzaam aan aan stimulus en handhaaft activiteit zolang de stimulus werd toegepast. Receptoren zijn verbonden met A? vezels en hebben een groot receptief veld. A3: Pacinische bloedlichaampjes zijn dieper in de dermis en detecteren trillingen. Receptoren zijn verbonden met A? vezels; ze passen zich snel aan stimulus aan en hebben het grootste receptieve veld. (B) Hele huid. B1: Merkel-celcomplexen zijn aanwezig in zowel de kale huid als rond het haar. Ze liggen dicht in de hand en zijn belangrijk voor textuurperceptie en het fijnste onderscheid tussen twee punten. Ze zijn verantwoordelijk voor de vingerprecisie. Receptoren zijn verbonden met A? vezels; ze passen zich langzaam aan aan stimulus en hebben een kort receptief veld. B2: Schadelijke aanraking HTMR's met zeer langzame aanpassing aan de stimulus, dwz actief zolang de nociceptieve stimulus wordt toegepast. Ze worden gevormd door het vrije zenuwuiteinde van A? en C-vezels geassocieerd met keratinocyten. (C) Harige huid. C1: Haarzakjes worden geassocieerd met de verschillende haartypes. Bij muizen zijn wachtharen het langste en spaarzaam tot expressie gebrachte, priem/auchenne zijn van gemiddelde grootte en zigzag zijn de kleinste en meest dichte haren. Ze zijn aangesloten op A? vezels, maar ook naar A? en C-LTMR-vezels voor priem/auchenne- en zizaghaar. Ze detecteren haarbewegingen, inclusief een aangename aanraking tijdens het strelen. Ze passen zich snel of met tussenliggende kinetiek aan stimulus aan. C2: C-touch zenuwuiteinden komen overeen met een subtype van C-vezels terminus met vrij uiteinde gekenmerkt door een lage mechanische drempel. Ze worden verondersteld te coderen voor een aangenaam gevoel dat wordt veroorzaakt door streling. Ze passen zich matig aan aan stimulus en hebben een kort receptief veld. Vermeende mechanosensitieve (MS) ionkanalen die tot expressie worden gebracht in de verschillende tactiele receptoren, worden dienovereenkomstig aangegeven op basis van voorlopige gegevens en vatten de huidige hypothese samen die wordt geëvalueerd.
Een tweede belangrijke populatie die werd geïdentificeerd, betreft de A?-LTMR-uiteinden in Awl/Auchenne en zigzag-follikels die moeten worden vergeleken met de donshaarfollikel die uitgebreid is bestudeerd bij katten en konijnen. Ginty en medewerkers toonden aan dat TrkB op hoge niveaus tot expressie wordt gebracht in een subset van DRG-neuronen met gemiddelde diameter. Intracellulaire opnames met behulp van de ex vivo huid-zenuwbereiding van gelabelde vezels onthulden dat ze de fysiologische eigenschappen vertonen van vezels die eerder zijn bestudeerd bij katten en konijnen: uitstekende mechanische gevoeligheid (Von Frey-drempel < 0.07 mN), snelle aanpassing van reacties op supradrempelstimuli, intermediaire geleiding snelheden (5.8 - 0.9 m/s) en smalle onverbogen soma spikes.15 Deze A?-LTMR's vormen longitudinale lancetvormige uiteinden die geassocieerd zijn met vrijwel elke zigzag- en priem/auchene haarfollikel van de romp [Fig. 1 (A5)].
Ten slotte toonden ze aan dat de perifere uitgangen van snel aanpassende A? LTMR's vormen longitudinale lancetvormige uiteinden geassocieerd met guard (of tylotrich) en priem/auchene haarzakjes [Fig. 1 (A6)].15 Bovendien worden bewakingsharen ook geassocieerd met een Merkel-celcomplex dat een aanraakkoepel vormt die is verbonden met A? langzaam aanpassen van LTMR [Fig. 1 (A7)].
Samengevat worden vrijwel alle zigzaghaarzakjes geïnnerveerd door zowel C-LTMR als A?-LTMR lancetvormige uiteinden; priem/auchene haren worden driemaal geïnnerveerd door A? snel aanpassende LTMR, A?-LTMR en C-LTMR lancetvormige uiteinden; Guard haarzakjes worden geïnnerveerd door A? snel aanpassen-LTMR longitudinale lancetvormige uiteinden en interactie met A? langzaam aanpassen-LTMR van touch dome-uiteinden. Zo ontvangt elke muishaarzakje unieke en onveranderlijke combinaties van LTMR-uiteinden die overeenkomen met neurofysiologisch verschillende mechanosensorische eindorganen. Gezien de iteratieve rangschikking van deze drie haartypes, stellen Ginty en medewerkers voor dat een harige huid bestaat uit een iteratieve herhaling van een perifere eenheid met (1) een of twee centraal geplaatste dekharen, (2) ~20 omringende priem-/auchenne-haren en (3 ) ~80 afgewisselde zigzagharen [Fig. 2 (C1)].
Projectie van het ruggenmerg. De centrale projecties van A? snel aanpassende LTMR's, AP-LTMR's en C-LTMR's eindigen in verschillende, maar gedeeltelijk overlappende laminae (II, III, IV) van de dorsale hoorn van het ruggenmerg. Bovendien zijn de centrale uiteinden van LTMR's die dezelfde of aangrenzende haarzakjes binnen een perifere LTMR-eenheid innerveren, uitgelijnd om een smalle LTMR-kolom te vormen in de dorsale hoorn van het ruggenmerg [Fig. 1 (B1)]. Het lijkt dus waarschijnlijk dat een wig of kolom van somatotopisch georganiseerde primaire sensorische afferente uiteinden in de dorsale hoorn de uitlijning vertegenwoordigt van de centrale projecties van A?-, A?- en C-LTMR's die dezelfde perifere eenheid innerveren en mechanische detecteren. prikkels die op dezelfde kleine groep haarzakjes inwerken. Op basis van het aantal bewakers, priem- en zigzagharen van de romp en ledematen en het aantal van elk LTMR-subtype, schatten Ginty en medewerkers dat de dorsale hoorn van de muis 2,000-4,000 LTMR-kolommen bevat, wat overeenkomt met het geschatte aantal perifere LTMR-eenheden.15
Bovendien zijn axonen van LTMR-subtypes nauw met elkaar verbonden, met verstrengelde uitsteeksels en in elkaar grijpende lancetvormige uiteinden die dezelfde haarfollikel innerveren. Omdat de drie soorten haarzakjes verschillende vormen, maten en celsamenstellingen vertonen, is het bovendien waarschijnlijk dat ze verschillende afbuigende of vibrerende afstemmingseigenschappen hebben. Deze bevindingen komen overeen met klassieke neurofysiologische metingen bij de kat en het konijn die erop wijzen dat A? RA-LTMR's en A?-LTMR's kunnen differentieel worden geactiveerd door afbuiging van verschillende haarzakjes.16,17
Concluderend, aanraking in een harige huid is de combinatie van: (1) de relatieve aantallen, unieke ruimtelijke verdelingen en duidelijke morfologische en afbuigende eigenschappen van de drie soorten haarzakjes; (2) de unieke combinaties van LTMR-subtype-uiteinden geassocieerd met elk van de drie haarfollikeltypes; en (3) verschillende gevoeligheden, geleidingssnelheden, spike-treinpatronen en aanpassingseigenschappen van de vier hoofdklassen van haarfollikel-geassocieerde LTMR's die het mechanosensorische systeem van de harige huid in staat stellen om de complexe combinaties van eigenschappen die een aanraken.
Vrije zenuwuiteinden LTMR's
Over het algemeen zijn C-vezels vrije uiteinden in de huid HTMR's, maar een subpopulatie van C-vezels reageert niet op schadelijke aanraking. Deze subset van tactiele C-vezel (CT) afferenten vertegenwoordigt een duidelijk type niet-gemyeliniseerde, laagdrempelige mechanoreceptieve eenheden die voorkomen in de harige maar niet-glabreuze huid van mensen en zoogdieren [Fig. 1 (A8)].18,19 CT's worden over het algemeen geassocieerd met de perceptie van aangename tactiele stimulatie bij lichaamscontact.20,21
CT-afferenten reageren op indrukkrachten in het bereik van 0.3-2.5 mN en zijn dus net zo gevoelig voor huidvervorming als veel van de A? afferenten.19 De aanpassingskenmerken van CT-afferenten zijn dus intermediair in vergelijking met de langzaam en snel aanpassende gemyeliniseerde mechanoreceptoren. De receptieve velden van menselijke CT-afferenten zijn ruwweg rond of ovaal van vorm. Het veld bestaat uit één tot negen kleine responsieve plekken verdeeld over een gebied tot 35 mm2.22 De homologe receptoren van de muis zijn georganiseerd in een patroon van onderbroken vlekken die ongeveer 50-60% van het gebied in de harige huid beslaan [Fig. 2 (C2)].23
Bewijs van patiënten zonder gemyeliniseerde tactiele afferenten geeft aan dat signalering in CT-vezels de insulaire cortex activeert. Aangezien dit systeem slecht is in het coderen van onderscheidende aspecten van aanraking, maar goed geschikt is voor het coderen van langzame, zachte aanraking, kunnen CT-vezels in een harige huid deel uitmaken van een systeem voor het verwerken van aangename en sociaal relevante aspecten van aanraking.24 CT-vezelactivering kan ook spelen een rol bij pijnremming en recentelijk is gesuggereerd dat ontsteking of trauma het gevoel dat wordt overgebracht door C-fiber LTMR's kan veranderen van aangename aanraking in pijn.25,26
Welke route CT-afferenten afleggen is nog niet bekend [Fig. 1 (B2)], maar laagdrempelige tactiele inputs voor spinothalamische projectiecellen zijn gedocumenteerd,27 wat geloofwaardigheid verleent aan meldingen van subtiele, contralaterale tekortkomingen van aanrakingsdetectie bij menselijke patiënten na vernietiging van deze paden na chordotomieprocedures.28
LTMR's in een gladde huid
Merkel cel-neuriet complexen en touch dome. Merkel (1875) was de eerste die een histologische beschrijving gaf van clusters van epidermale cellen met grote gelobde kernen, die contact maken met veronderstelde afferente zenuwvezels. Hij nam aan dat ze de tastzin onderhielden door ze Tastzellen (tactiele cellen) te noemen. Bij mensen zijn Merkel-cel-neurietcomplexen verrijkt in aanraakgevoelige delen van de huid, ze worden aangetroffen in de basale laag van de opperhuid in vingers, lippen en geslachtsorganen. Ze komen ook voor in een harige huid met een lagere dichtheid. Het Merkel-cel-neurietcomplex bestaat uit een Merkel-cel in nauwe appositie aan een vergrote zenuwuiteinde van een enkele gemyeliniseerde A? vezel [afb. 1 (C1)] (overzicht in Halata en medewerkers).29 Aan de epidermale zijde vertoont de Merkel-cel vingerachtige processen die zich uitstrekken tussen naburige keratinocyten [Fig. 1 (C2)]. Merkelcellen zijn van keratinocyten afgeleide epidermale cellen.30,31 De term touch dome werd geïntroduceerd om de grote concentratie van Merkel-celcomplexen in de harige huid van de voorpoot van de kat te noemen. Een aanraakkoepel kan tot 150 Merkel-cellen bevatten die worden geïnnerveerd door een enkele A?-vezel en bij mensen, naast A?-vezels, A? en C-vezels waren ook regelmatig aanwezig.32-34
Stimulatie van Merkel-cel-neurietcomplexen resulteert in langzaam aanpassende Type I (SA I) reacties, die afkomstig zijn van puntige receptieve velden met scherpe randen. Er is geen spontane ontlading. Deze complexen reageren op de inkepingsdiepte van de huid en hebben de hoogste ruimtelijke resolutie (0.5 mm) van de cutane mechanoreceptoren. Ze zenden een nauwkeurig ruimtelijk beeld van tactiele stimuli uit en er wordt aangenomen dat ze verantwoordelijk zijn voor vorm- en textuurdiscriminatie [Fig. 2 (B1)]. Muizen zonder Merkel-cellen kunnen met hun voeten geen gestructureerde oppervlakken detecteren terwijl ze dit met hun snorharen doen
Of de Merkel-cel, het sensorische neuron of beide plaatsen van mechanotransductie zijn, is nog steeds een kwestie van debat. Bij ratten heft de fototoxische vernietiging van Merkel-cellen de SA I-respons op.36 Bij muizen met genetisch onderdrukte Merkel-cellen verdween de SA I-respons die werd geregistreerd in ex vivo huid/zenuwpreparaat volledig, wat aantoont dat Merkel-cellen nodig zijn voor de juiste codering van Merkel receptorreacties.37 De mechanische stimulatie van geïsoleerde Merkel-cellen in kweek door motoraangedreven druk genereert echter geen mechanisch gestuurde stromen.38,39 Keratinocyten kunnen een belangrijke rol spelen bij de normale werking van het Merkel-cel-neurietcomplex. De vingerachtige processen van de Merkelcel kunnen bewegen met huidvervorming en epidermiscelbeweging, en dit kan de eerste stap zijn van mechanische transductie. Het is duidelijk dat de voorwaarden die nodig zijn om de mechanische gevoeligheid van Merkel-cellen te bestuderen, nog moeten worden vastgesteld.
Ruffini eindes. Ruffini-uiteinden zijn dunne sigaarvormige ingekapselde sensorische uiteinden die zijn verbonden met A? zenuwuiteinden. Ruffini-uiteinden zijn kleine bindweefselcilinders die zijn gerangschikt langs dermale collageenstrengen die worden geleverd door één tot drie gemyeliniseerde zenuwvezels met een diameter van 4 m. Maximaal drie cilinders met verschillende oriëntatie in de dermis kunnen samensmelten om één receptor te vormen [Fig. 6 (C1)]. Structureel zijn Ruffini-uiteinden vergelijkbaar met Golgi-peesorganen. Ze worden algemeen uitgedrukt in de dermis en zijn geïdentificeerd als de langzaam aanpassende type II (SA II) cutane mechanoreceptoren. Tegen de achtergrond van spontane zenuwactiviteit wordt een langzaam aanpassende regelmatige ontlading opgewekt door loodrechte mechanische stimulatie met lage kracht of effectiever door dermale rek. SA II-respons is afkomstig van grote receptieve velden met onduidelijke grenzen. Ruffini-receptoren dragen bij aan de perceptie van de richting van objectbeweging door het patroon van huidrek [Fig. 3 (A2)].
Bij muizen kunnen SA I- en SA II-responsen elektrofysiologisch worden gescheiden in ex-vivo zenuwhuidbereiding.40 Nandasena en medewerkers rapporteerden de immunolokalisatie van aquaporine 1 (AQP1) in de parodontale Ruffini-uiteinden van de snijtanden van de rat, wat suggereert dat AQP1 betrokken is bij het handhaven van de tandheelkundige osmotische balans die nodig is voor de mechanotransductie.41 De parodontale Ruffini-uiteinden drukten ook het vermeende mechanosensitieve ionkanaal uit ASIC3.42
bloedlichaampjes van Meissner. De bloedlichaampjes van Meissner zijn gelokaliseerd in de dermale papillen van de kale huid, voornamelijk in de handpalmen en voetzolen, maar ook in de lippen, in de tong, in het gezicht, in de tepels en in de geslachtsorganen. Anatomisch gezien bestaan ze uit een ingekapseld zenuwuiteinde, waarbij het kapsel is gemaakt van afgeplatte ondersteunende cellen die zijn gerangschikt als horizontale lamellen die zijn ingebed in bindweefsel. Er is één enkele zenuwvezel A? afferenten verbonden per bloedlichaampje [Fig. 1 (C4)]. Elke fysieke vervorming van het lichaampje veroorzaakt een stroom van actiepotentialen die snel ophoudt, dat wil zeggen, ze zijn snel aanpassende receptoren. Wanneer de stimulus wordt verwijderd, krijgt het bloedlichaampje zijn vorm terug en produceert het een nieuw salvo van actiepotentialen. Vanwege hun oppervlakkige locatie in de dermis reageren deze bloedlichaampjes selectief op huidbewegingen, tactiele detectie van uitglijden en trillingen (20-40 Hz). Ze zijn gevoelig voor dynamische huid – bijvoorbeeld tussen de huid en een voorwerp dat wordt gehanteerd [Fig. 2 (A1)].
Pacinische bloedlichaampjes. Pacinische bloedlichaampjes zijn de diepere mechanoreceptoren van de huid en zijn de meest gevoelige ingekapselde huidmechanoreceptor van huidbeweging. Deze grote eivormige bloedlichaampjes (1 mm lang) gemaakt van concentrische lamellen van vezelig bindweefsel en fibroblasten omzoomd door platte gemodificeerde Schwann-cellen worden uitgedrukt in de diepe dermis.43 In het midden van het lichaampje, in een met vloeistof gevulde holte genaamd binnenste bol , beëindigt een enkele A? afferente niet-gemyeliniseerde zenuwuiteinde [Fig. 1 (C5)]. Ze hebben een groot receptief veld op het huidoppervlak met een bijzonder gevoelig centrum. De ontwikkeling en functie van verschillende snel aanpassende mechanoreceptortypen zijn verstoord in c-Maf-mutante muizen. In het bijzonder zijn Pacinische bloedlichaampjes ernstig geatrofieerd
Pacinische bloedlichaampjes vertonen een zeer snelle aanpassing in reactie op de inkeping van de huid, de snel aanpassende II (RA II) zenuwontlading die in staat is om hoogfrequente trillingsstimuli te volgen en waarneming van gebeurtenissen op afstand mogelijk te maken door uitgezonden trillingen.45 Pacinische bloedlichaampjes afferenten reageren op aanhoudende inspringing met voorbijgaande activiteit bij het begin en de verschuiving van de stimulus. Ze worden ook versnellingsdetectoren genoemd omdat ze veranderingen in de sterkte van de stimulus kunnen detecteren en, als de snelheid van verandering in de stimulus wordt gewijzigd (zoals gebeurt bij trillingen), wordt hun reactie evenredig met deze verandering. De bloedlichaampjes van Pacinian nemen grote drukveranderingen waar en vooral trillingen (150-300 Hz), die ze zelfs op centimeters afstand kunnen detecteren [Fig. 2 (A3)].
Tonische respons werd waargenomen in ontkapselde Pacinische bloedlichaampjes.46 Bovendien reageren intacte Pacinische bloedlichaampjes met aanhoudende activiteit tijdens constante inspringstimuli, zonder mechanische drempels of responsfrequentie te veranderen wanneer GABA-gemedieerde signalering wordt geblokkeerd tussen gelamelleerde glia en een zenuwuiteinde.47 de niet-neuronale componenten van het Pacinische bloedlichaampje kunnen een dubbele rol spelen bij het filteren van de mechanische stimulus en bij het moduleren van de responseigenschappen van het sensorische neuron.
Ruggenmerg projecties. Projecties van de A?-LTMR's in het ruggenmerg zijn verdeeld in twee takken. De belangrijkste centrale tak stijgt in het ruggenmerg in de ipsilaterale dorsale kolommen naar het cervicale niveau [Fig. 1 (B3)]. Secundaire takken eindigen in de dorsale hoorn in de laminae IV en interfereren bijvoorbeeld met de pijnoverdracht. Dit kan pijn verminderen als onderdeel van de poortcontrole [Fig. 1 (B4)].48
Op cervicaal niveau scheiden axonen van de hoofdtak zich in twee delen: het middellijnkanaal omvat de gracile bundel die informatie van de onderste helft van het lichaam (benen en romp) transporteert en het buitenste kanaal omvat de cuneate bundel die informatie van de bovenste helft transporteert van het lichaam (armen en romp) [Fig. 1 (B5)].
Primaire tactiele afferenten maken hun eerste synaps met neuronen van de tweede orde in de medulla waar vezels van elk kanaal synapsen in een kern met dezelfde naam: de gracile fasciculus-axonen synaps in de gracile kern en de cuneate axonen synaps in de cuneate-kern [Fig. 1 (B6)]. Neuronen die de synaps ontvangen, leveren de secundaire afferenten en steken onmiddellijk de middellijn over om een kanaal te vormen aan de contralaterale zijde van de hersenstam - de mediale lemniscus - die opstijgt door de hersenstam naar het volgende relaisstation in de middenhersenen, in het bijzonder in de thalamus [Fig. . 1 (B7)].
Moleculaire specificatie van LTMR's. Moleculaire mechanismen die de vroege diversificatie van LTMR's beheersen, zijn onlangs gedeeltelijk opgehelderd. Bourane en medewerkers hebben aangetoond dat de neuronale populaties die de Ret-tyrosinekinasereceptor (Ret) en zijn co-receptor GFR?2 tot expressie brengen in E11-13 embryonale muizen DRG selectief de transcriptiefactor Mafa tot co-expressie brengen.49,50 Deze auteurs tonen aan dat de Mafa/ Ret/GFR?2-neuronen zijn voorbestemd om bij de geboorte drie specifieke typen LTRM's te worden: de SA1-neuronen die Merkel-celcomplexen innerveren, de snel aanpassende neuronen die de Meissner-bloedlichaampjes innerveren en de snel aanpassende afferenten (RA I) die lancetvormige uiteinden rond de haarzakjes vormen. Ginty en medewerkers melden ook dat DRG-neuronen die vroeg-Ret tot expressie brengen, snel de mechanoreceptoren van Meissner-bloedlichaampjes, Pacinische bloedlichaampjes en lancetvormige uiteinden rond haarzakjes aanpassen.51 Ze innerveren afzonderlijke doelzones binnen de gracile en cuneate kernen, wat een modaliteit-specifiek patroon van mechanosensorische neuron axonale projecties in de hersenstam.
Onderzoek naar mechanoreceptoren van de menselijke huid. De techniek van "microneurografie" beschreven door Hagbarth en Vallbo in 1968 is toegepast om het ontladingsgedrag te bestuderen van enkelvoudige menselijke mechanosensitieve uiteinden die spieren, gewrichten en huid voeden (zie voor een overzicht Macefield, 2005).52,53 Het merendeel van de menselijke huidmicroneurografie studies hebben de fysiologie van tactiele afferenten in de kale huid van de hand gekarakteriseerd. Micro-elektrode-opnames van de mediane en ulnaire zenuwen bij menselijke proefpersonen hebben een aanrakingsgevoel onthuld dat wordt gegenereerd door de vier klassen van LTMR's: Meissner-afferenten zijn bijzonder gevoelig voor licht dat over de huid strijkt, reagerend op lokale schuifkrachten en beginnende of openlijke slips binnen het receptieve veld. Pacinische afferenten zijn buitengewoon gevoelig voor snelle mechanische transiënten. Afferenten reageren heftig op het blazen over het receptieve veld. Een Pacinisch lichaampje dat zich in een cijfer bevindt, reageert meestal op het tikken op de tafel die de arm ondersteunt. Merkel-afferenten hebben kenmerkend een hoge dynamische gevoeligheid voor inkepingsstimuli die op een afzonderlijk gebied worden toegepast en reageren vaak met een uit-ontlading tijdens het loslaten. Hoewel de afferenten van Ruffini reageren op krachten die normaal op de huid worden uitgeoefend, is een uniek kenmerk van SA II-afferenten hun vermogen om ook te reageren op laterale huidrek. Ten slotte hebben haareenheden in de onderarm grote eivormige of onregelmatige receptieve velden die zijn samengesteld uit meerdere gevoelige plekken die overeenkomen met individuele haren (elk afferente levert ~20 haren).
Mechanische gevoeligheid van keratinocyten
Elke mechanische stimulus op de huid moet worden overgedragen via keratinocyten die de epidermis vormen. Deze alomtegenwoordige cellen kunnen naast hun ondersteunende of beschermende rol ook signaleringsfuncties vervullen. Keratinocyten scheiden bijvoorbeeld ATP af, een belangrijk sensorisch signaalmolecuul, als reactie op mechanische en osmotische stimuli.54,55 De afgifte van ATP induceert intracellulaire calciumtoename door autocriene stimulatie van purinerge receptoren.55 Verder zijn er aanwijzingen dat hypotonie de Rho activeert. -kinase-signaleringsroute en de daaropvolgende vorming van F-actine-stressvezels, wat suggereert dat de mechanische vervorming van de keratinocyten mechanisch kan interfereren met de naburige cellen zoals Merkel-cellen voor onschadelijke aanraking en C-vezelvrije uiteinden voor schadelijke aanraking [Fig. 1 (C6)].56,57
Schadelijke aanraking
Mechanoreceptoren met hoge drempel (HTMR's) zijn epidermale C- en A? vrije zenuwuiteinden. Ze zijn niet geassocieerd met gespecialiseerde structuren en worden waargenomen in zowel de harige huid [Fig. 1 (A9)] en een kale huid [Fig. 1(C7)]. De term vrije zenuwuiteinden moet echter voorzichtig worden overwogen, aangezien zenuwuiteinden altijd dicht bij elkaar liggen met keratinocyten of Langherans-cellen of melanocyten. Ultrastructurele analyse van zenuwuiteinden onthult de aanwezigheid van ruw endoplasmatisch reticulum, overvloedige mitochondriën en blaasjes met een dichte kern. Aangrenzende membranen van epidermale cellen zijn verdikt en lijken op postsynaptische membraan in zenuwweefsel. Merk op dat de interacties tussen zenuwuiteinden en epidermale cellen bidirectioneel kunnen zijn, aangezien epidermale cellen mediatoren zoals ATP, interleukine (IL6, IL10) en bradykinine kunnen afgeven en omgekeerd kunnen peptiderge zenuwuiteinden peptiden zoals CGRP of substantie P afgeven die op epidermale cellen inwerken. HTMR's omvatten mechano-nociceptoren die alleen worden opgewonden door schadelijke mechanische stimuli en polymodale nociceptoren die ook reageren op schadelijke hitte en exogene chemicaliën [Fig. 2 (B2)].58
HTMR afferente vezels eindigen op projectie-neuronen in de dorsale hoorn van het ruggenmerg. A?-HTMR's maken voornamelijk contact met neuronen van de tweede orde in de lamina I en V, terwijl C-HTMR's eindigen in de lamina II [Fig. 1 (B8)]. Tweede-orde nociceptieve neuronen projecteren naar de controlaterale zijde van het ruggenmerg en stijgen op in de witte stof, en vormen het anterolaterale systeem. Deze neuronen eindigen voornamelijk in de thalamus [Fig. 1 (B9 en B10)].
Mechano-stromen in somatosensorische neuronen
De mechanismen van langzame of snelle aanpassing van mechanoreceptoren zijn nog niet opgehelderd. Het is niet duidelijk in welke mate mechanoreceptoraanpassing wordt geleverd door de cellulaire omgeving van het sensorische zenuwuiteinde, de intrinsieke eigenschappen van de mechanisch-gated kanalen en de eigenschappen van de axonale spanningsafhankelijke ionkanalen in sensorische neuronen (Fig. 2). Recente vooruitgang in de karakterisering van mechanisch gereguleerde stromen heeft echter aangetoond dat er verschillende klassen van mechanosensitieve kanalen bestaan in DRG-neuronen en sommige aspecten van de aanpassing van mechanoreceptoren kunnen verklaren.
In vitro-opnames bij knaagdieren hebben aangetoond dat het soma van DRG-neuronen intrinsiek mechanogevoelig is en kationische mechano-gated stromen tot expressie brengt.59-64 Gadolinium en rutheniumrood blokkeren mechanisch gevoelige stromen volledig, terwijl extern calcium en magnesium, bij fysiologische concentraties, evenals amiloride en benzamil, veroorzaken een gedeeltelijke blokkering.60,62,63 FM1-43 werkt als een blijvende blokker, en de injectie van FM1-43 in de achterpoot van muizen vermindert de pijngevoeligheid in de Randall-Selitto-test en verhoogt de beoordeelde drempel voor het terugtrekken van de poot met von Frey haren.65
Als reactie op aanhoudende mechanische stimulatie nemen mechanosensitieve stromen af door sluiting. Op basis van de tijdconstanten van stroomafname zijn vier verschillende soorten mechanisch gevoelige stromen onderscheiden: snel aanpassende stromen (~3 ms), tussentijds aanpassende stromen (~6 ms), langzaam aanpassende stromen (~15-30 ms) ) en ultra-traag aanpassende stromen (~200 ms).300 Al deze stromen zijn aanwezig met variabele incidentie in DRG-neuronen van ratten die de kale huid van de achterpoot innerveren.1000
De mechanische gevoeligheid van mechanisch gevoelige stromen kan worden bepaald door een reeks incrementele mechanische stimuli toe te passen, waardoor een relatief gedetailleerde stimulus-stroomanalyse mogelijk is.66 De stimulus-stroomrelatie is typisch sigmoïdaal en de maximale amplitude van de stroom wordt bepaald door het aantal kanalen die tegelijkertijd open zijn.64,67 Interessant is dat de snel aanpassende mechanisch gevoelige stroom een lage mechanische drempel en een half activeringsmiddelpunt vertoont in vergelijking met de ultra langzaam aanpassende mechanisch gevoelige stroom.63,65
Sensorische neuronen met niet-nociceptieve fenotypes brengen bij voorkeur snel aanpassende mechanosensitieve stromen tot expressie met een lagere mechanische drempel.60,61,63,64,68 Omgekeerd worden langzaam en ultra-langzaam aanpassende mechanosensitieve stromen af en toe gerapporteerd in vermeende niet-nociceptieve cellen.64,68 Dit leidde tot de suggestie dat deze stromen zouden kunnen bijdragen aan de verschillende mechanische drempels die worden gezien in LTMR's en HTMR's in vivo. Hoewel deze in vitro-experimenten met de nodige voorzichtigheid moeten worden uitgevoerd, werd ondersteuning voor de aanwezigheid in de soma van de DRG-neuronen van laag- en hoogdrempelige mechanotransducers ook geleverd door op radiale rek gebaseerde stimulatie van gekweekte sensorische neuronen van muizen.69 Dit paradigma onthulde twee belangrijkste populaties van rekgevoelige neuronen, een die reageert op een lage stimulusamplitude en een andere die selectief reageert op een hoge stimulusamplitude.
Deze resultaten hebben belangrijke, maar speculatieve, mechanistische implicaties: de mechanische drempel van sensorische neuronen heeft misschien weinig te maken met de cellulaire organisatie van de mechanoreceptor, maar kan liggen in de eigenschappen van de mechanisch gepoorte ionkanalen.
De mechanismen die ten grondslag liggen aan desensibilisatie van mechanosensitieve kationstromen in DRG-neuronen van ratten zijn onlangs ontrafeld.64,67 Het is het resultaat van twee gelijktijdige mechanismen die kanaaleigenschappen beïnvloeden: aanpassing en inactivatie. Aanpassing werd voor het eerst gemeld in auditieve haarcelstudies. Het kan operationeel worden beschreven als een eenvoudige vertaling van de activeringscurve van het transducerkanaal langs de mechanische stimulus-as.70-72 Aanpassing stelt sensorische receptoren in staat om hun gevoeligheid voor nieuwe stimuli te behouden in de aanwezigheid van een bestaande stimulus. Een aanzienlijk deel van de mechanisch gevoelige stromen in DRG-neuronen kan echter niet worden gereactiveerd na conditionerende mechanische stimulatie, wat wijst op inactivering van sommige transducerkanalen.64,67 Daarom werken zowel inactivatie als aanpassing samen om mechanisch gevoelige stromen te reguleren. Deze twee mechanismen zijn gemeenschappelijk voor alle mechanosensitieve stromen die zijn geïdentificeerd in DRG-neuronen van ratten, wat suggereert dat verwante fysisch-chemische elementen de kinetiek van deze kanalen bepalen.64
Concluderend is het bepalen van de eigenschappen van endogene mechanosensitieve stromen in vitro cruciaal in de zoektocht naar het identificeren van transductiemechanismen op moleculair niveau. De waargenomen variabiliteit in de mechanische drempel en de zich aanpassende kinetiek van de verschillende mechanisch gestuurde stromen in DRG-neuronen suggereren dat intrinsieke eigenschappen van ionenkanalen, ten minste gedeeltelijk, de mechanische drempel en aanpassingskinetiek van de mechanoreceptoren beschreven in de decennia 1960 kunnen verklaren� 80 met behulp van ex vivo preparaten.
Vermeende mechanisch gevoelige eiwitten
Mechanosensitieve ionenstromen in somatosensorische neuronen zijn goed gekarakteriseerd, er is daarentegen weinig bekend over de identiteit van moleculen die mechanotransductie bij zoogdieren bemiddelen. Genetische screenings in Drosophila en C. elegans hebben kandidaat-mechanotransductiemoleculen geïdentificeerd, waaronder de TRP- en degenerine/epitheliale Na+-kanaal (Deg/ENaC) families.73 Recente pogingen om de moleculaire basis van mechanotransductie bij zoogdieren op te helderen, waren grotendeels gericht op homologen van deze kandidaten . Bovendien zijn veel van deze kandidaten aanwezig in cutane mechanoreceptoren en somatosensorische neuronen (Fig. 2).
Zuurgevoelige ionenkanalen
ASIC's behoren tot een proton-gated subgroep van de degeneratie-epitheliale Na+-kanaalfamilie.74 Drie leden van de ASIC-familie (ASIC1, ASIC2 en ASIC3) komen tot expressie in mechanoreceptoren en nociceptoren. De rol van ASIC-kanalen is onderzocht in gedragsstudies met muizen met gerichte verwijdering van ASIC-kanaalgenen. Deletie van ASIC1 verandert de functie van cutane mechanoreceptoren niet, maar verhoogt de mechanische gevoeligheid van afferenten die de darmen innerveren.75 ASIC2-knockout-muizen vertonen een verminderde gevoeligheid van zich snel aanpassende cutane LTMR's.76 Latere studies meldden echter een gebrek aan effecten van het uitschakelen van ASIC2 op zowel viscerale mechano-nociceptie als cutane mechanosensatie.77 ASIC3-verstoring vermindert de mechanische gevoeligheid van viscerale afferenten en vermindert de respons van cutane HTMR's op schadelijke stimuli.76
Het voorbijgaande receptorkanaal
DE TRP-superfamilie is onderverdeeld in zes subfamilies bij zoogdieren.78 Bijna alle TRP-subfamilies hebben leden die verband houden met mechanosensatie in een verscheidenheid aan celsystemen.79 In sensorische neuronen van zoogdieren zijn TRP-kanalen echter het best bekend voor het detecteren van thermische informatie en het mediëren van neurogene ontstekingen. en slechts twee TRP-kanalen, TRPV4 en TRPA1, zijn betrokken bij de aanraakreactie. Het verstoren van TRPV4-expressie bij muizen heeft slechts bescheiden effecten op acute mechanosensorische drempels, maar vermindert de gevoeligheid voor schadelijke mechanische stimuli sterk.80,81 TRPV4 is een cruciale determinant bij het vormgeven van de respons van nociceptieve neuronen op osmotische stress en op mechanische hyperalgesie tijdens ontsteking.82,83, 1 TRPA1 lijkt een rol te spelen bij mechanische hyperalgesie. TRPA1-deficiënte muizen vertonen overgevoeligheid voor pijn. TRPA84,85 draagt bij aan de transductie van mechanische, koude en chemische stimuli in nociceptor sensorische neuronen, maar het blijkt niet essentieel te zijn voor haarceltransductie.XNUMX
Er is geen duidelijk bewijs dat aangeeft dat TRP-kanalen en ASIC-kanalen die tot expressie worden gebracht in zoogdieren mechanisch zijn gepoort. Geen van deze kanalen die heteroloog tot expressie zijn gebracht, recapituleert de elektrische signatuur van mechanisch gevoelige stromen waargenomen in hun oorspronkelijke omgeving. Dit sluit de mogelijkheid niet uit dat ASIC's en TRP-kanalen mechanotransducers zijn, gezien de onzekerheid of een mechanotransductiekanaal buiten zijn cellulaire context kan functioneren (zie de sectie over SLP3).
Piëzo-eiwitten
Piëzo-eiwitten zijn recentelijk geïdentificeerd als veelbelovende kandidaten voor mechanosensing-eiwitten door Coste en medewerkers.86,87 Gewervelde dieren hebben twee piëzo-leden, Piezo 1 en Piezo 2, voorheen bekend als respectievelijk FAM38A en FAM38B, die goed geconserveerd zijn in meercellige eukaryoten . Piezo 2 is overvloedig aanwezig in DRG's, terwijl Piezo 1 nauwelijks detecteerbaar is. Piëzo-geïnduceerde mechanisch gevoelige stromen worden voorkomen, geremd door gadolinium, ruthenium rood en GsMTx4 (een toxine van de tarantula Grammostola spatulata).88 Expressie van piëzo 1 of piëzo 2 in heterologe systemen produceert mechanosensitieve stromen, de kinetiek van inactivering van piëzo 2-stroom is sneller dan piëzo 1. Net als bij endogene mechanisch gevoelige stromen, hebben piëzo-afhankelijke stromen omkeerpotentialen rond 0 mV en zijn ze kation-niet-selectief, waarbij Na+, K+, Ca2+ en Mg2+ allemaal het onderliggende kanaal doordringen. Evenzo worden piëzo-afhankelijke stromen gereguleerd door membraanpotentiaal, met een duidelijke vertraging van de stroomkinetiek bij gedepolariseerde potentialen.86
Piëzo-eiwitten zijn ongetwijfeld mechanosensing-eiwitten en delen veel eigenschappen van snel aanpassende mechanosensitieve stromen in sensorische neuronen. Behandeling van gekweekte DRG-neuronen met piëzo 2 kort interfererend RNA verminderde het aandeel neuronen met snel aanpassende stroom en verlaagde het percentage mechanosensitieve neuronen.86 Transmembraandomeinen bevinden zich door de piëzo-eiwitten, maar er zijn geen duidelijke poriebevattende motieven of ionkanaalsignaturen gevonden. geïdentificeerd. Piezo 1-eiwit van muis is echter gezuiverd en gereconstitueerd in asymmetrische lipidedubbellagen en liposoomvormen ionkanalen die gevoelig zijn voor rutheniumrood.87 Een essentiële stap bij het valideren van mechanotransductie via piëzo-kanalen is het gebruik van in vivo benaderingen om het functionele belang bij aanraaksignalering te bepalen. Er werd informatie gegeven in Drosophila waar deletie van het enkele piëzo-lid de mechanische respons op schadelijke stimuli verminderde, zonder de normale aanraking te beïnvloeden.89 Hoewel hun structuur nog moet worden bepaald, is deze nieuwe familie van mechanosensitieve eiwitten een veelbelovend onderwerp voor toekomstig onderzoek, over de grens van aanrakingssensatie. Een recent onderzoek bij patiënten met anemie (erfelijke xerocytose) toont bijvoorbeeld de rol van Piezo 1 bij het handhaven van de homeostase van het erytrocytenvolume.90
Transmembraan kanaalachtig (TMC)
Een recente studie geeft aan dat twee eiwitten, TMC1 en TMC2, nodig zijn voor mechanotransductie van haarcellen.91 Erfelijke doofheid als gevolg van TMC1-genmutatie is gemeld bij mens en muis.92,93 Aanwezigheid van deze kanalen was nog niet aangetoond in het somatosensorische systeem , maar het lijkt een goede aanleiding om te onderzoeken.
Stomatine-achtige proteïne 3 (SLP3)
Naast de transductiekanalen is aangetoond dat sommige accessoire-eiwitten die aan het kanaal zijn gekoppeld, een rol spelen bij de aanrakingsgevoeligheid. SLP3 wordt tot expressie gebracht in DRG-neuronen van zoogdieren. Studies met gemuteerde muizen zonder SLP3 hadden verandering in mechanosensatie en mechanosentive stromen aangetoond.94,95 De precieze functie van SLP3 blijft onbekend. Het kan een schakel zijn tussen het mechanisch gevoelige kanaal en de onderliggende microtubuli, zoals voorgesteld voor zijn C. elegans homoloog MEC2.96 Recent GR. Lewin Lab heeft gesuggereerd dat een ketting wordt gesynthetiseerd door DRG-sensorische neuronen en het mechanosensitieve ionkanaal koppelt aan de extracellulaire matrix.97 Het verstoren van de link heft de RA-mechanosensitieve stroom op, wat suggereert dat sommige ionenkanalen alleen mechanosensitief zijn wanneer ze zijn vastgemaakt. RA-mechanosensitieve stromen worden ook geremd door laminine-332, een matrixeiwit geproduceerd door keratinocyten, wat de hypothese van een modulatie van de mechanosensitieve stroom door extracellulaire eiwitten versterkt.98
K+ kanaal subfamilie
Parallel aan kationische depolariserende mechanosensitieve stromen wordt de aanwezigheid van repolariserende mechanosensitieve K+ stromen onderzocht. K+-kanalen in mechanosensitieve cellen kunnen in de huidige balans stappen en bijdragen aan het bepalen van de mechanische drempel en het tijdsverloop van de aanpassing van mechanoreceptoren.
KCNK-leden behoren tot de familie van het K+-kanaal (K2P) met twee poriën.99,100 De K2P vertonen een opmerkelijk bereik van regulatie door cellulaire, fysieke en farmacologische middelen, waaronder pH-veranderingen, hitte, rek en membraanvervorming. Deze K2P zijn actief bij rustmembraanpotentiaal. Verschillende KCNK-subeenheden worden tot expressie gebracht in somatosensorische neuronen.101 KCNK2 (TREK-1), KCNK4 (TRAAK) en TREK-2 kanalen behoren tot de weinige kanalen waarvoor een directe mechanische poorting door membraanrek is aangetoond.102,103
Muizen met een verstoord KCNK2-gen vertoonden een verhoogde gevoeligheid voor warmte en milde mechanische stimuli, maar een normale terugtrekkingsdrempel voor schadelijke mechanische druk uitgeoefend op de achterpoot met behulp van de Randall-Selitto-test.104 KCNK2-deficiënte muizen vertonen ook verhoogde thermische en mechanische hyperalgesie bij inflammatoire conditie. KCNK4-knockout-muizen waren overgevoelig voor milde mechanische stimulatie en deze overgevoeligheid werd verhoogd door extra inactivering van KCNK2.105. Verhoogde mechanosensitiviteit van deze knock-outmuizen zou kunnen betekenen dat rek normaal zowel depolariserende als repolariserende mechanosensitieve stromen op een gecoördineerde manier activeert, vergelijkbaar met de onbalans van depolariserende en repolariserende spanningsafhankelijke stromen.
KCNK18 (TRESK) levert een belangrijke bijdrage aan de achtergrond K+-geleiding die de rustmembraanpotentiaal van somatosensorische neuronen reguleert.106 Hoewel het niet bekend is of KCNK18 direct gevoelig is voor mechanische stimulatie, kan het een rol spelen bij het mediëren van reacties op lichte aanraking, evenals pijnlijke mechanische stimuli. KCNK18 en in mindere mate KCNK3 worden voorgesteld als het moleculaire doelwit van hydroxy-?-sanshool, een verbinding die wordt aangetroffen in Schezuan-peperkorrels die aanraakreceptoren activeert en een tintelend gevoel bij mensen veroorzaakt.107,108
Het spanningsafhankelijke K+-kanaal KCNQ4 (Kv7.4) is cruciaal voor het instellen van de snelheids- en frequentievoorkeur van een subpopulatie van zich snel aanpassende mechanoreceptoren bij zowel muizen als mensen. Mutatie van KCNQ4 werd aanvankelijk in verband gebracht met een vorm van erfelijke doofheid. Interessant is dat een recente studie KCNQ4 lokaliseert in de perifere zenuwuiteinden van de huid, zich snel aanpassend haarzakje en Meissner-lichaampje. Dienovereenkomstig leidt verlies van KCNQ4-functie tot een selectieve verbetering van de gevoeligheid van de mechanoreceptor voor laagfrequente trillingen. Met name mensen met laat optredend gehoorverlies als gevolg van dominante mutaties van het KCNQ4-gen vertonen verbeterde prestaties bij het detecteren van trillingen met een kleine amplitude en een lage frequentie.109

Insight van Dr. Alex Jimenez
Aanraking wordt beschouwd als een van de meest complexe zintuigen in het menselijk lichaam, vooral omdat er geen specifiek orgaan voor is. In plaats daarvan vindt de tastzin plaats via sensorische receptoren, ook wel mechanoreceptoren genoemd, die over de huid worden aangetroffen en reageren op mechanische druk of vervorming. Er zijn vier hoofdtypen mechanoreceptoren in de kale of haarloze huid van zoogdieren: lamellaire bloedlichaampjes, tactiele bloedlichaampjes, Merkel-zenuwuiteinden en bolvormige bloedlichaampjes. Mechanoreceptoren werken om de detectie van aanraking mogelijk te maken, om de positie van de spieren, botten en gewrichten te bewaken, bekend als proprioceptie, en zelfs om geluiden en de beweging van het lichaam te detecteren. Het begrijpen van de mechanismen van structuur en functie van deze mechanoreceptoren is een fundamenteel element in het gebruik van behandelingen en therapieën voor pijnbestrijding.
Conclusie
Aanraking is een complex zintuig omdat het verschillende tactiele kwaliteiten vertegenwoordigt, namelijk vibratie, vorm, textuur, plezier en pijn, met verschillende onderscheidende prestaties. Tot nu toe was de overeenkomst tussen een tastorgaan en de psychofysieke zin correlatief en klasse-specifieke moleculaire markers zijn net in opkomst. De ontwikkeling van knaagdiertests die overeenkomen met de diversiteit van aanrakingsgedrag is nu vereist om toekomstige genomics-identificatie te vergemakkelijken. Het gebruik van muizen die specifieke subsets van sensorische afferente typen missen, zal de identificatie van mechanoreceptoren en sensorische afferente vezels die verband houden met een bepaalde aanrakingsmodaliteit aanzienlijk vergemakkelijken. Interessant is dat een recent artikel de belangrijke vraag opent naar de genetische basis van mechanosensorische eigenschappen bij de mens en suggereert dat een enkele genmutatie de aanrakingsgevoeligheid negatief zou kunnen beïnvloeden.110 Dit onderstreept dat de pathofysiologie van het menselijke aanrakingstekort voor een groot deel onbekend is en zeker vooruitgang door precies de subset van sensorische neuronen te identificeren die verband houden met een aanrakingsmodaliteit of een aanrakingstekort.
In ruil daarvoor is er vooruitgang geboekt bij het definiëren van de biofysische eigenschappen van de mechanisch gestuurde stromen.64 De ontwikkeling van nieuwe technieken in de afgelopen jaren, die het mogelijk maken om veranderingen in de membraanspanning te volgen, terwijl de mechanisch gestuurde stroom wordt geregistreerd, is een waardevolle experimentele methode gebleken om de mechanosensitieve stromen met snelle, intermediaire en langzame aanpassing (besproken in Delmas en medewerkers).66,111 De toekomst zal zijn om de rol van de huidige eigenschappen in de mechanismen van aanpassing van functioneel diverse mechanoreceptoren en de bijdrage van mechanosensitieve K+ stromen aan de prikkelbaarheid van LTMR's en HTMR's.
De moleculaire aard van mechanisch gestuurde stromen bij zoogdieren is ook een toekomstig veelbelovend onderzoeksonderwerp. Toekomstig onderzoek zal plaatsvinden in twee perspectieven, eerst om de rol te bepalen van een accessoire-molecuul dat kanalen aan het cytoskelet bindt en dat nodig zou zijn om mechanosensitiviteit van ionenkanalen zoals TRP- en ASIC/EnaC-families te verlenen of te reguleren. Ten tweede, om het grote en veelbelovende gebied van de bijdrage van de piëzo-kanalen te onderzoeken door sleutelvragen te beantwoorden, met betrekking tot de permeatie- en poortmechanismen, de subset van sensorische neuronen en aanrakingsmodaliteiten waarbij piëzo betrokken is en de rol van piëzo in niet-neuronale cellen geassocieerd met mechanosensatie.
De tastzin kan, in vergelijking met die van zicht, smaak, geluid en geur, die specifieke organen gebruiken om deze gewaarwordingen te verwerken, door het hele lichaam voorkomen via kleine receptoren die bekend staan als mechanoreceptoren. Verschillende soorten mechanoreceptoren zijn te vinden in verschillende lagen van de huid, waar ze een breed scala aan mechanische stimulatie kunnen detecteren. Het artikel hierboven beschrijft specifieke hoogtepunten die de voortgang aantonen van structurele en functionele mechanismen van mechanoreceptoren die verband houden met de tastzin. Informatie waarnaar wordt verwezen door het National Center for Biotechnology Information (NCBI). De reikwijdte van onze informatie is beperkt tot zowel chiropractie als letsels en aandoeningen van de wervelkolom. Als u het onderwerp wilt bespreken, kunt u het aan Dr. Jimenez vragen of contact met ons opnemen via�915-850-0900 .
Curator van Dr. Alex Jimenez

Bijkomende onderwerpen: Rugpijn
Rugpijn is een van de meest voorkomende oorzaken van invaliditeit en gemiste dagen op het werk wereldwijd. Rugpijn is trouwens toegeschreven als de op één na meest voorkomende reden voor bezoeken aan artsenbureaus, die in de minderheid zijn dan alleen infecties aan de bovenste luchtwegen. Ongeveer 80 procent van de bevolking ervaart ten minste één keer gedurende hun hele leven een vorm van rugpijn. De wervelkolom is een complexe structuur bestaande uit botten, gewrichten, ligamenten en spieren, naast andere zachte weefsels. Vanwege dit, verwondingen en / of verergerde omstandigheden, zoals herniated discs, kan uiteindelijk leiden tot symptomen van rugpijn. Sportblessures of ongevallen met auto-ongelukken zijn vaak de meest voorkomende oorzaak van rugpijn, maar soms kunnen de eenvoudigste bewegingen pijnlijke resultaten hebben. Gelukkig kunnen alternatieve behandelingsopties, zoals chiropractische zorg, de rugpijn verlichten door het gebruik van spinale aanpassingen en handmatige manipulaties, waardoor uiteindelijk de pijnverlichting wordt verbeterd.

EXTRA BELANGRIJK ONDERWERP: Beheer van lage rugpijn
MEER ONDERWERPEN: EXTRA EXTRA: Chronische pijn en behandelingen
Blanco
Referenties
Accordeon sluiten
Professionele reikwijdte van de praktijk *
De informatie hierin over "Structurele en functionele mechanismen van mechanoreceptoren" is niet bedoeld ter vervanging van een een-op-een relatie met een gekwalificeerde zorgverlener of gediplomeerde arts en is geen medisch advies. We moedigen u aan om beslissingen over de gezondheidszorg te nemen op basis van uw onderzoek en samenwerking met een gekwalificeerde zorgverlener.
Blog Informatie & Scope Discussies
Ons informatiebereik: is beperkt tot chiropractie, musculoskeletale, fysieke medicijnen, welzijn, bijdragende etiologische viscerosomatische stoornissen binnen klinische presentaties, geassocieerde somatoviscerale reflex klinische dynamiek, subluxatiecomplexen, gevoelige gezondheidskwesties en/of functionele geneeskunde artikelen, onderwerpen en discussies.
Wij bieden en presenteren klinische samenwerking met specialisten uit verschillende disciplines. Elke specialist wordt beheerst door hun professionele praktijk en hun jurisdictie van licentiestatus. We gebruiken functionele gezondheids- en welzijnsprotocollen om de verwondingen of aandoeningen van het bewegingsapparaat te behandelen en te ondersteunen.
Onze video's, berichten, onderwerpen, onderwerpen en inzichten behandelen klinische zaken, problemen en onderwerpen die verband houden met en direct of indirect onze klinische praktijk ondersteunen.*
Ons kantoor heeft redelijkerwijs geprobeerd om ondersteunende citaten te geven en heeft de relevante onderzoeksstudie of studies geïdentificeerd die onze berichten ondersteunen. Wij verstrekken kopieën van ondersteunende onderzoeksstudies die op verzoek beschikbaar zijn voor regelgevende instanties en het publiek.
We begrijpen dat we zaken behandelen die een aanvullende uitleg vereisen over hoe het kan helpen bij een bepaald zorgplan of behandelprotocol; daarom, om het onderwerp hierboven verder te bespreken, aarzel dan niet om te vragen Dr. Alex Jimenez, DC, of neem contact met ons op 915-850-0900.
Wij zijn er om u en uw gezin te helpen.
zegeningen
Dr. Alex Jimenez DC MSACP, RN*, CCST, IFMCP*, CIFM*, ATN*
e-mail: coach@elpasofunctionalmedicine.com
Licentie als Doctor of Chiropractic (DC) in Texas & New Mexico*
Texas DC-licentie # TX5807, New Mexico DC-licentie # NM-DC2182
Licentie als geregistreerde verpleegkundige (RN*) in Florida
Florida-licentie RN-licentie # RN9617241 (controle nr. 3558029)
Compacte status: Licentie voor meerdere staten: Geautoriseerd om te oefenen in 40 Staten*
Dr. Alex Jimenez DC, MSACP, RN* CIFM*, IFMCP*, ATN*, CCST
Mijn digitale visitekaartje





