Back Clinic Oxidatieve Stress Chiropractie en Functionele Geneeskunde Team. Oxidatieve stress wordt gedefinieerd als een verstoring in de balans tussen de productie van reactieve zuurstof (vrije radicalen) en de afweer van antioxidanten. Met andere woorden, het is een disbalans tussen de productie van vrije radicalen en het vermogen van het lichaam om de schadelijke effecten tegen te gaan of te ontgiften door neutralisatie door antioxidanten. Oxidatieve stress leidt tot veel pathofysiologische aandoeningen in het lichaam. Deze omvatten neurodegeneratieve ziekten, dwz de ziekte van Parkinson, de ziekte van Alzheimer, genmutaties, kankers, chronisch vermoeidheidssyndroom, fragiel X-syndroom, hart- en bloedvataandoeningen, atherosclerose, hartfalen, hartaanval en ontstekingsziekten. Oxidatie vindt plaats onder een aantal omstandigheden:
de cellen gebruiken glucose om energie te maken
het immuunsysteem bestrijdt bacteriën en veroorzaakt ontstekingen
de lichamen ontgiften verontreinigende stoffen, pesticiden en sigarettenrook
Er zijn miljoenen processen op elk moment in ons lichaam die kunnen leiden tot oxidatie. Hier zijn een paar symptomen:
Vermoeidheid
Geheugenverlies en of hersenmist
Spier- en / of gewrichtspijn
Rimpels samen met grijs haar
Verminderd gezichtsvermogen
Hoofdpijn en gevoeligheid voor lawaai
Gevoeligheid voor infecties
Het kiezen van biologisch voedsel en het vermijden van gifstoffen in uw omgeving maakt een groot verschil. Dit, samen met het verminderen van stress, kan gunstig zijn bij het verminderen van oxidatie.
Oxidanten worden over het algemeen op een gecontroleerde manier geproduceerd om essentiële processen in het menselijk lichaam te reguleren, waaronder celdeling, ontsteking, immuunfunctie, autofagie en stressrespons. De ongecontroleerde productie van deze oxidanten kan echter bijdragen oxidatieve stress, die de cellulaire functie kunnen beïnvloeden, wat leidt tot de ontwikkeling van toxiciteit, chronische ziekten en kanker. De beschermende antioxidantmechanismen van het menselijk lichaam worden gereguleerd door een reeks vitale routes die de reactie van de cel op oxidanten regelen. De nucleaire factor erytroïde 2-gerelateerde factor, ook wel Nrf2 genoemd, is een opkomende regulator van cellulaire weerstand tegen oxidanten. Het doel van het onderstaande artikel is om de opkomende rol van Nrf2 in de mitochondriale functie te bespreken en aan te tonen.
Abstract
De transcriptiefactor NF-E2 p45-gerelateerde factor 2 (Nrf2; gennaam NFE2L2) maakt aanpassing en overleving onder stressomstandigheden mogelijk door de genexpressie van diverse netwerken van cytoprotectieve eiwitten te reguleren, waaronder ook antioxidant-, ontstekingsremmende en ontgiftingsenzymen als eiwitten die helpen bij het herstel of de verwijdering van beschadigde macromoleculen. Nrf2 speelt een cruciale rol bij het in stand houden van cellulaire redoxhomeostase door de biosynthese, het gebruik en de regeneratie van glutathion, thioredoxine en NADPH te reguleren en door de productie van reactieve zuurstofsoorten door mitochondriën en NADPH-oxidase te regelen. Onder homeostatische omstandigheden beïnvloedt Nrf2 het mitochondriale membraanpotentieel, vetzuuroxidatie, beschikbaarheid van substraten (NADH en FADH2 / succinaat) voor ademhaling en ATP-synthese. Onder omstandigheden van stress of groeifactorstimulatie, gaat activering van Nrf2 de verhoogde productie van reactieve zuurstofsoorten in mitochondriën tegen via transcriptionele opregulatie van ontkoppelingseiwit 3 en beïnvloedt mitochondriale biogenese door de niveaus van nucleaire ademhalingsfactor 1 en peroxisoom-proliferator-geactiveerde receptor te handhaven? coactivator 1?, evenals door het bevorderen van de biosynthese van purine-nucleotiden. Farmacologische Nrf2-activators, zoals het natuurlijk voorkomende isothiocyanaat sulforafaan, remmen door oxidatiemiddel gemedieerde opening van de mitochondriale permeabiliteitstransitieporie en mitochondriale zwelling. Vreemd genoeg bleek een synthetische 1,4-difenyl-1,2,3-triazoolverbinding, oorspronkelijk ontworpen als een Nrf2-activator, mitofagie te bevorderen en daarmee bij te dragen aan de algehele mitochondriale homeostase. Nrf2 is dus een prominente speler in het ondersteunen van de structurele en functionele integriteit van de mitochondriën, en deze rol is vooral cruciaal onder stressomstandigheden.
Nrf2 speelt een cruciale rol bij het handhaven van cellulaire redox-homeostase.
Nrf2 beïnvloedt de mitochondriale membraanpotentiaal en ATP-synthese.
Nrf2 beïnvloedt de mitochondriale vetzuuroxidatie.
Nrf2 ondersteunt de structurele en functionele integriteit van de mitochondriën.
Nrf2-activators hebben gunstige effecten wanneer de mitochondriale functie wordt aangetast.
Introductie
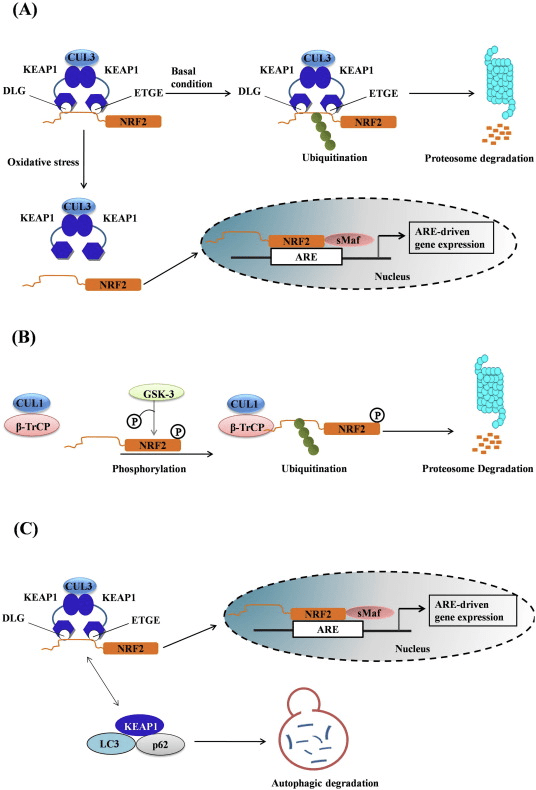
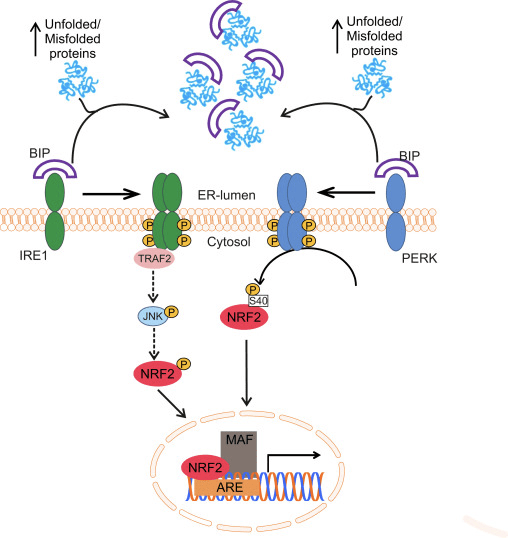
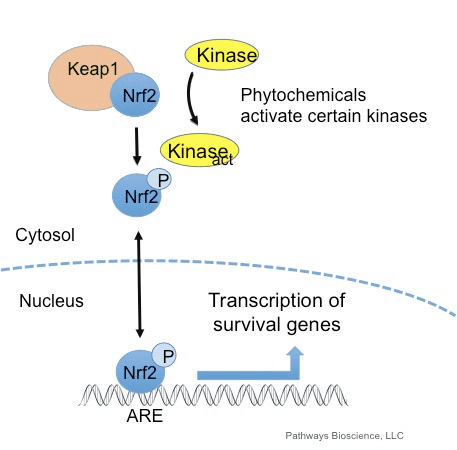
De transcriptiefactor NF-E2 p45-gerelateerde factor 2 (Nrf2; gennaam NFE2L2) reguleert de expressie van netwerken van genen die coderen voor eiwitten met diverse cytoprotectieve activiteiten. Nrf2 zelf wordt voornamelijk gecontroleerd op het niveau van eiwitstabiliteit. Onder basale omstandigheden is Nrf2 een kortlevend eiwit dat wordt onderworpen aan continue ubiquitinatie en proteasomale afbraak. Er zijn drie bekende ubiquitine-ligasesystemen die bijdragen aan de afbraak van Nrf2. Historisch gezien was de eerste negatieve regulator van Nrf2 die werd ontdekt Kelch-achtige ECH-geassocieerde proteïne 1 (Keap1) [1], een substraatadaptereiwit voor Cullin 3 (Cul3) / Rbx1 ubiquitine-ligase [2], [3], [ 4]. Keap1 gebruikt een zeer efficiënt cyclisch mechanisme om Nrf2 te richten voor ubiquitinatie en proteasomale afbraak, waarbij Keap1 continu wordt geregenereerd, waardoor de cyclus kan doorgaan (Fig. 1A) [5]. Nrf2 wordt ook onderworpen aan degradatie die wordt gemedieerd door glycogeensynthasekinase (GSK) 3 /? - TrCP-afhankelijke Cul1-gebaseerde ubiquitine-ligase [6], [7]. Recentelijk werd gerapporteerd dat Nrf2 tijdens condities van endoplasmatisch reticulumstress wordt ubiquitineerd en afgebroken in een proces dat wordt gemedieerd door het E3-ubiquitine-ligase Hrd1 [8].
Figuur 1 Het cyclische sequentiële bindings- en regeneratiemodel voor Keap1-gemedieerde afbraak van Nrf2. (A) Nrf2 bindt sequentieel aan een gratis Keap1-dimeer: eerst via het hoge-affiniteits-ETGE (rode staafjes) -bindende domein en vervolgens via het lage-affiniteit DLG (zwarte staafjes) -bindende domein. In deze conformatie van het proteïnecomplex ondergaat Nrf2 ubiquitinatie en is het gericht op proteasomale afbraak. Gratis Keap1 wordt geregenereerd en kan binden aan nieuw vertaalde Nrf2, en de cyclus begint opnieuw. (B) Inductoren (witte ruiten) reageren met sensorcysteïnen van Keap1 (blauwe sticks), wat leidt tot een conformationele verandering en verminderde substraatadapteractiviteit. Gratis Keap1 wordt niet geregenereerd en de nieuw gesynthetiseerde Nrf2 accumuleert en verplaatst zich naar de kern.
Naast het dienen als een ubiquitine-ligase substraatadaptatie-eiwit, is Keap1 ook de sensor voor een breed scala aan kleine molecuul-activatoren van Nrf2 (aangeduid als inductoren) [9]. Inductoren blokkeren de cyclus van Keap1-gemedieerde afbraak van Nrf2 door chemische cysteïneresiduen in Keap1 [10], [11] chemisch te modificeren of door de Keap1: Nrf2-bindingsinterface [12], [13] rechtstreeks te verstoren. Dientengevolge wordt Nrf2 niet afgebroken, en de transcriptiefactor accumuleert en transloceert naar de kern (Fig. 1B), waar het een heterodimeer vormt met een klein Maf-eiwit; bindt aan antioxidant-responselementen, de stroomopwaartse regulatorische gebieden van zijn doelwitgenen; en initieert transcriptie [14], [15], [16]. De batterij van Nrf2-targets omvat eiwitten met diverse cytoprotectieve functies, waaronder enzymen van xenobiotisch metabolisme, eiwitten met antioxiderende en ontstekingsremmende functies en proteasomale subeenheden, evenals eiwitten die cellulaire redox-homeostase reguleren en deelnemen aan intermediair metabolisme.
Nrf2: een meester-regulator van cellulaire redox-homeostase
De functie van Nrf2 als hoofdregulator van cellulaire redoxhomeostase wordt algemeen erkend. De genexpressie van zowel de katalytische als de regulerende subeenheden van ?-glutamylcysteïneligase, het enzym dat de snelheidsbeperkende stap in de biosynthese van gereduceerd glutathion (GSH) katalyseert, wordt direct gereguleerd door Nrf2 [17]. De xCT-subeenheid van systeem xc-, die cystine in cellen importeert, is ook een direct transcriptioneel doelwit van Nrf2 [18]. In de cel wordt cystine omgezet in cysteïne, een voorloper voor de biosynthese van GSH. Naast zijn rol in GSH-biosynthese, biedt Nrf2 de middelen voor het behoud van glutathion in zijn gereduceerde staat door de gecoördineerde transcriptionele regulatie van glutathionreductase 1 [19], [20], die geoxideerd glutathion reduceert tot GSH met behulp van reducerende equivalenten van NADPH . De vereiste NADPH wordt geleverd door vier belangrijke NADPH-genererende enzymen, appelzuur 1 (ME1), isocitraatdehydrogenase 1 (IDH1), glucose-6-fosfaatdehydrogenase (G6PD) en 6-fosfogluconaatdehydrogenase (PGD), die allemaal transcriptioneel gereguleerd gedeeltelijk door Nrf2 (Fig. 2) [21], [22], [23], [24]. Vreemd genoeg reguleert Nrf2 ook de induceerbare genexpressie van de cytosolische, microsomale en mitochondriale vormen van aldehydedehydrogenase [25], die NAD(P)+ als cofactor gebruiken, waardoor NAD(P)H ontstaat. Inderdaad, de niveaus van NADPH en de NADPH / NADP + -verhouding zijn lager in embryonale fibroblasten die zijn geïsoleerd uit Nrf2-knockout (Nrf2-KO) muizen in vergelijking met cellen van hun wildtype (WT) tegenhangers, en de NADPH-niveaus nemen af na Nrf2 knockdown in kankercellijnen met constitutief actieve Nrf2 [26]. Zoals verwacht zijn de niveaus van GSH lager in cellen waarin Nrf2 is verstoord; omgekeerd leidt Nrf2-activering door genetische of farmacologische middelen tot GSH-upregulatie [27], [28], [29]. Belangrijk is dat Nrf2 ook de genexpressie reguleert van thioredoxine [30], [31], [32], thioredoxine-reductase 1 [28], [29], [32], [33] en sulfidoxine [34], die essentieel zijn voor de reductie van geoxideerde eiwitthiolen.
Figuur 2 De rol van Nrf2 in het metabolisme van snel prolifererende cellen. Nrf2 is een positieve regulator van genen die coderen voor enzymen in zowel de oxidatieve arm [dwz glucose-6-fosfaat dehydrogenase (G6PD) en 6-fosfogluconaat dehydrogenase (PGD)] en de niet-oxidatieve arm [dwz transaldolase 1 (TALDO1) en transketolase ( TKT)] van de pentosefosfaatroute. G6PD en PGD genereren NADPH. Nrf2 reguleert ook de genexpressie van de andere twee NADPH-genererende enzymen, appelzuur-enzym 1 (ME1) en isocitraat dehydrogenase 1 (IDH1). De genexpressie van fosforibosylpyrofosfaatamidotransferase (PPAT), die de toegang tot de de novo purine biosynthetische route katalyseert, wordt ook positief gereguleerd door Nrf2, evenals de expressie van methyleentetrahydrofolaat dehydrogenase 2 (MTHFD2), een mitochondriaal enzym met een cruciale rol in het leveren van eenheden van één koolstof voor de novo purinebiosynthese. Pyruvaatkinase (PK) wordt negatief gereguleerd door Nrf2 en zal naar verwachting de opbouw van glycolytische tussenproducten bevorderen en, samen met G6PD, metabolietkanalen door de pentosefosfaatroute en de synthese van nucleïnezuren, aminozuren en fosfolipiden. Nrf2 reguleert negatief de genexpressie van ATP-citraatlyase (CL), wat de beschikbaarheid van citraat voor mitochondriaal gebruik of (via isocitraat) voor IDH1 kan verhogen. Rood en blauw geven respectievelijk positieve en negatieve regulering aan. Het mitochondrion wordt grijs weergegeven. Afkortingen voor metabolieten: G-6-P, glucose-6-fosfaat; F-6-P, fructose 6-fosfaat; F-1,6-BP, fructose 1,6-bisfosfaat; GA-3-P, glyceraldehyde 3-fosfaat; 3-PG, 3-fosfoglyceraat; PEP, fosfoenolpyruvaat; 6-P-Gl, 6-fosfogluconolacton; 6-PG, 6-fosfogluconaat; R-5-P, ribulose 5-fosfaat; PRPP, 5-fosforibosyl - a - 1-pyrofosfaat; THF, tetrahydrofolaat; IMP, inosine monofosfaat; AMP, adenosinemonofosfaat; GMP, guanosinemonofosfaat.
Gezien de cruciale rol van Nrf2 als een meesterregulator van cellulaire redox-homeostase, is het niet verrassend dat, in vergelijking met WT-cellen, de niveaus van reactieve zuurstofspecies (ROS) hoger zijn in cellen waarin Nrf2 is verstoord (Nrf2-KO) [35]. Dit verschil is vooral opvallend bij een aanval met middelen die oxidatieve stress veroorzaken. Bovendien zijn cellen die deficiënt zijn in Nrf2 veel gevoeliger voor de toxiciteit van oxidanten van verschillende typen en kunnen ze niet worden beschermd door Nrf2-induceerders, die onder dezelfde omstandigheden efficiënte en langdurige bescherming bieden voor WT-cellen [29], [36] , [37]. Naast de algemene cellulaire redox-homeostase is Nrf2 ook van cruciaal belang voor het behoud van de mitochondriale redox-homeostase. In vergelijking met WT is de totale mitochondriale NADH-pool dus aanzienlijk verhoogd in Keap1-KO en dramatisch gedaald in Nrf2-KO-cellen [35].
Met behulp van live cell imaging hebben we onlangs de snelheden van de ROS-productie in primaire glioneurale co-culturen en hersenweefselplakjes geïsoleerd uit WT-, Nrf2-KO- of Keap1-knockdown-muizen (Keap1-KD) [38] gemonitord. Zoals verwacht, was de snelheid van de ROS-productie in de Nrf2-KO-cellen en -weefsels hoger in vergelijking met hun WT-tegenhangers. We hebben echter de onverwachte waarneming gemaakt dat, in vergelijking met WT, Keap1-KD-cellen ook hogere ROS-productiecijfers hebben, hoewel de grootte van het verschil tussen de WT- en de Keap1-KD-genotypes kleiner was dan die tussen WT en Nrf2-KO . Vervolgens analyseerden we de mRNA-niveaus van NOX2 en NOX4, de katalytische subeenheden van de twee NADPH-oxidase (NOX) -isovormen die betrokken zijn bij hersenpathologie, en vonden dat NOX2 dramatisch is verhoogd onder omstandigheden van Nrf2-tekort, terwijl NOX4 opgereguleerd wordt wanneer Nrf2 is constitutief geactiveerd, zij het in mindere mate. Kwantitatief loopt de grootte van opregulatie in cellen en weefsels van de mutante muizen parallel met de overeenkomstige toenames in ROS-productie [38]. Interessant is dat nietfxNUMX niet alleen NADPH-oxidase reguleert, maar dat de ROS geproduceerd door NADPH-oxidase Nrf2 kan activeren, zoals aangetoond in pulmonale epitheliale cellen en cardiomyocyten [2], [39]. Bovendien heeft een zeer recente studie aangetoond dat de NADPH-oxidase-afhankelijke activering van Nrf40 een belangrijk endogeen mechanisme vormt voor bescherming tegen mitochondriale schade en celdood in het hart tijdens chronische overbelasting van de druk [2].
Naast de katalytische activiteit van NADPH-oxidase, is mitochondriale ademhaling een andere belangrijke intracellulaire bron van ROS. Met behulp van de mitochondriën-specifieke probe MitoSOX hebben we de bijdrage van ROS van mitochondriale oorsprong aan de totale ROS-productie in primaire glioneurale co-culturen onderzocht. van WT, Nrf2-KO of Keap1-KD-muizen [38]. Zoals verwacht hadden Nrf2-KO-cellen hogere percentages mitochondriale ROS-productie dan WT. In overeenstemming met de bevindingen voor de totale ROS-productie waren de percentages van mitochondriale ROS-productie in Keap1-KD ook hoger in vergelijking met WT-cellen. Belangrijk is dat het blokkeren van complex I met rotenon een dramatische toename van de mitochondriale ROS-productie in zowel WT- als Keap1-KD-cellen veroorzaakte, maar geen effect had in Nrf2-KO-cellen. In tegenstelling tot de verwachte toename in mitochondriale ROS-productie in WT-cellen na toevoeging van pyruvaat (ter verbetering van de beschikbaarheid van NADH, verhoging van het mitochondriaal membraanpotentieel en normalisering van de ademhaling), nam de productie van ROS af in Nrf2-KO-cellen. Samen suggereren deze bevindingen sterk dat, in de afwezigheid van Nrf2: (i) de activiteit van complex I is verminderd, (ii) de verminderde activiteit van complex I te wijten is aan beperking van substraten, en (iii) de verminderde activiteit van complex Ik is een van de belangrijkste redenen voor de verhoogde mitochondriale ROS-productie, mogelijk als gevolg van de omgekeerde elektronenstroom uit complex II.
Nrf2 beïnvloedt mitochondriaal membraanpotentiaal en ademhaling
De mitochondriale membraanpotentiaal (?? m) is een universele indicator van de mitochondriale gezondheid en de metabolische toestand van de cel. In een gezonde cel wordt ?? m in stand gehouden door de mitochondriale ademhalingsketen. Interessant is dat een stabiele isotopische labeling met aminozuren in op kweek gebaseerde proteomics-studie in de oestrogeenreceptor-negatieve niet-tumorigene MCF10A-cellijn van de menselijke borstepitheel heeft aangetoond dat de mitochondriale elektronentransportketencomponent NDUFA4 wordt opgereguleerd door farmacologische activering (door sulforafaan) van Nrf2, terwijl genetische opregulatie van Nrf2 (door Keap1 knockdown) leidt tot neerwaartse regulatie van de cytochroom c oxidase subeenheden COX2 en COX4I1 [42]. Een studie van het leverproteoom met behulp van tweedimensionale gelelektroforese en matrix-geassisteerde laserdesorptie / ionisatie massaspectrometrie heeft aangetoond dat Nrf2 de expressie van ATP-synthase-subeenheid reguleert? [43]. Bovendien is gemeld dat het mitochondriale eiwit DJ-1, dat een rol speelt bij het behoud van de activiteit van complex I [44], Nrf2 [45], [46] stabiliseert, hoewel de neuroprotectieve effecten van farmacologische of genetische activering van Nrf2 zijn onafhankelijk van DJ-1 [47]. De gevolgen van deze waarnemingen voor de mitochondriale functie zijn echter niet onderzocht.
In overeenstemming met de verminderde activiteit van complex I onder omstandigheden van Nrf2-deficiëntie, is de basale ?? m lager in Nrf2-KO-muizenembryofibroblasten (MEF's) en gekweekte primaire glioneuronale cellen in vergelijking met hun WT-tegenhangers (Fig. 3, inzet) [35]. Daarentegen is de basale ?? m hoger wanneer Nrf2 genetisch constitutief wordt opgereguleerd (door knock-out of knock-out van Keap1). Deze verschillen in ?? m tussen de genotypen geven aan dat de ademhaling wordt beïnvloed door de activiteit van Nrf2. Evaluatie van het zuurstofverbruik in de basale toestand heeft inderdaad aangetoond dat, vergeleken met WT, het zuurstofverbruik lager is in Nrf2-KO en Keap1-KO MEF's, met respectievelijk ~ 50 en ~ 35%.
Figuur 3 Voorgesteld mechanisme voor aangetaste mitochondriale functie onder omstandigheden van Nrf2-tekort. (1) De verlaagde niveaus van ME1, IDH1, G6PD en PGD resulteren in lagere NADPH-niveaus. (2) De GSH-niveaus zijn ook laag. (3) De lage activiteit van ME1 kan de pool van pyruvaat die de mitochondriën binnendringt verminderen. (4) De generatie van NADH is langzamer, wat leidt tot verminderde activiteit van complex I en verhoogde mitochondriale ROS-productie. (5) De reductie van FAD tot FADH2 in mitochondriale eiwitten is ook verminderd, waardoor de elektronenstroom van FADH2 naar UbQ en naar complex III wordt verlaagd. (6) De langzamere vorming van UbQH2 kan de enzymactiviteit van succinaatdehydrogenase verlagen. (7) De verhoogde ROS-niveaus kunnen de activiteit van complex II verder remmen. (8) De lagere efficiëntie van vetzuuroxidatie draagt bij aan de verminderde beschikbaarheid van substraat voor mitochondriale ademhaling. (9) Glycolyse wordt versterkt als een compensatiemechanisme voor de verminderde ATP-productie bij oxidatieve fosforylering. (10) ATP-synthase werkt omgekeerd om ?? m te behouden. Rood en blauw geven respectievelijk opregulatie en neerwaartse regulatie aan. De kaders duiden de beschikbaarheid van experimenteel bewijs aan. De inzet toont afbeeldingen van mitochondriën van WT en Nrf2-KO corticale astrocyten gevisualiseerd door de potentiometrische fluorescerende sonde tetramethylrhodamine methylester (TMRM; 25 nM). Schaalbalk, 20 m.
Deze verschillen in ?? m en ademhaling tussen de genotypen worden weerspiegeld door de mate van gebruik van substraten voor mitochondriale ademhaling. Toepassing van substraten voor de tricarbonzuur (TCA) cyclus (malaat / pyruvaat, die op hun beurt de productie van het complex I substraat NADH verhogen) of methylsuccinaat, een substraat voor complex II, zorgt voor een stapsgewijze toename van ?? m in beide WT en Keap1-KD-neuronen, maar de snelheid van toename is hoger in Keap1-KD-cellen. Wat nog belangrijker is, is dat de vormen van de respons op deze TCA-cyclussubstraten verschillen tussen de twee genotypen, waarbij de snelle stijging in μm in Keap1-KD-cellen na toevoeging van substraat wordt gevolgd door een snelle daling in plaats van een plateau, wat een ongebruikelijk snel substraatverbruik. Deze bevindingen zijn in nauwe overeenstemming met de veel lagere (met 50-70%) niveaus van malaat, pyruvaat en succinaat die zijn waargenomen na een puls van 1 uur van [U-13C6] glucose in Keap1-KO vergeleken met WT MEF cellen [24]. In Nrf2-KO-neuronen kan alleen pyruvaat de ?? m verhogen, terwijl malaat en methylsuccinaat milde depolarisatie veroorzaken. Het effect van Nrf2 op de productie van mitochondriaal substraat lijkt het belangrijkste mechanisme te zijn waarmee Nrf2 de mitochondriale functie beïnvloedt. De mitochondriale NADH-redoxindex (de balans tussen consumptie van NADH door complex I en productie van NADPH in de TCA-cyclus) is significant lager in Nrf2-KO-cellen in vergelijking met hun WT-tegenhangers, en bovendien is de regeneratiesnelheid van de pools van NADH en FADH2 na remming van complex IV (door gebruik van NaCN) zijn langzamer in de mutante cellen.
In mitochondriën geïsoleerd uit muizenhersenen en lever, verhoogt suppletie van substraten voor complex I of voor complex II de snelheid van zuurstofconsumptie sterker wanneer Nrf2 wordt geactiveerd en minder efficiënt wanneer Nrf2 wordt verstoord [35]. Malaat induceert dus een hoger zuurstofverbruik in Keap1-KD in vergelijking met WT, maar het effect is zwakker in Nrf2-KO-mitochondriën. Evenzo activeert succinaat in de aanwezigheid van rotenon (wanneer complex I wordt geremd) het zuurstofverbruik in grotere mate in Keap1-KD in vergelijking met WT, terwijl de respons in Nrf2-KO-mitochondriën verminderd is. Bovendien zijn Nrf2-KO primaire neuronale culturen en muizen gevoeliger voor de toxiciteit van de complex II-remmers 3-nitropropionzuur en malonaat, terwijl intrastriatale transplantatie van Nrf2-tot overexpressie brengende astrocyten beschermend is [48], [49]. Evenzo zijn Nrf2-KO-muizen gevoeliger voor, terwijl genetische of farmacologische activering van Nrf2 beschermende effecten heeft tegen neurotoxiciteit veroorzaakt door de complex I-remmer 1-methyl-4-fenylpyridiniumion in het 1-methyl-4-fenyl-1,2,3,6, 49-tetrahydropyridine diermodel van de ziekte van Parkinson [50], [51], [52], [53], [54], [55], [56], [57], [58], [59], [60], [61], [XNUMX].
De respiratory control ratio (RCR), de verhouding van State 3 (ADP-gestimuleerd) tot State 4-ademhaling (geen ADP aanwezig), is verlaagd in afwezigheid van Nrf2, maar de RCR is vergelijkbaar tussen Keap1-KD- en WT-mitochondriën [35 ]. Aangezien de RCR een indicatie is van de mate van koppeling van de mitochondriale ademhalingsketenactiviteit aan oxidatieve fosforylering, geeft deze bevinding aan dat de hogere ademhalingssnelheid in Keap1-KD-mitochondriën niet het gevolg is van ontkoppeling van oxidatieve fosforylering. Het suggereert verder dat oxidatieve fosforylering efficiënter is wanneer Nrf2 wordt geactiveerd. De hogere ademhalingssnelheid in Keap1-KD-mitochondriën is consistent met de hogere niveaus van mitochondriale ROS-productie [38], aangezien hogere ademhalingsfrequenties kunnen leiden tot een verhoogd elektronenlek. Onder omstandigheden van oxidatieve stress wordt de verhoogde ROS-productie echter tegengegaan door de Nrf2-afhankelijke transcriptionele opregulatie van ontkoppelingsproteïne 3 (UCP3), die de protongeleiding van het mitochondriale binnenmembraan verhoogt en bijgevolg de productie van superoxide verlaagt [62]. Zeer recent werd aangetoond dat het lipideperoxidatieproduct 4-hydroxy-2-nonenal de Nrf2-afhankelijke opregulatie van UCP3 in hartspiercellen medieert; dit kan met name belangrijk zijn voor bescherming onder omstandigheden van oxidatieve stress, zoals die tijdens ischemie reperfusie [63].
Nrf2 beïnvloedt de efficiëntie van oxidatieve fosforylering en de synthese van ATP
In overeenstemming met het effect van Nrf2 op de ademhaling, in hersen- en lever mitochondria, resulteert Nrf2-deficiëntie in een verminderde efficiëntie van oxidatieve fosforylering (zoals geschat door de verhouding van ADP tot zuurstof, die wordt verbruikt voor ATP-synthese), terwijl Nrf2-activering (Keap1 -KD) heeft het tegenovergestelde effect [35]. In vergelijking met WT zijn de ATP-niveaus significant hoger in cellen met constitutieve opregulatie van Nrf2 en lager wanneer Nrf2 wordt uitgeschakeld [64] of verstoord [35]. Bovendien heeft het gebruik van remmers van oxidatieve fosforylering (oligomycine) of glycolyse (joodazijnzuur) onthuld dat Nrf2 de manier verandert waarop cellen ATP produceren. Dus in WT-neuronen veroorzaakt oligomycine een volledige daling van ATP en heeft joodazijnzuur geen verder effect. Opmerkelijk is dat oligomycine in Nrf2-KO-cellen de ATP-niveaus verhoogt, die vervolgens langzaam maar volledig worden uitgeput door joodazijnzuur, wat aangeeft dat in de afwezigheid van Nrf2 glycolyse en niet oxidatieve fosforylering de belangrijkste bron van ATP-productie is. Interessant is dat, ondanks de verhoogde efficiëntie van oxidatieve fosforylering in Keap1-KD-cellen, toevoeging van oligomycine resulteert in een afname van ~ 80% in ATP-niveaus en dat joodazijnzuur een verdere afname van ~ 20% veroorzaakt. Dus ofwel Nrf2-deficiëntie of de constitutieve activering ervan vermindert de bijdrage van oxidatieve fosforylering en verhoogt de bijdrage van glycolyse aan de synthese van ATP. Dit effect is vooral uitgesproken wanneer Nrf2 afwezig is en is consistent met de afhankelijkheid van de ?? m van de aanwezigheid van glucose in het medium [35] en de verhoogde niveaus van glycolytische tussenproducten (G-6-P, F-6-P dihydroxyacetonfosfaat, pyruvaat en lactaat) na uitschakelen van Nrf2 [24].
De toename van ATP-niveaus na remming van de F1F0-ATPase door oligomycine geeft aan dat in de afwezigheid van Nrf2 de F1F0-ATPase functioneert als een ATPase en niet als een ATP-synthase, dwz het werkt in omgekeerde richting. Een dergelijke omkering in activiteit weerspiegelt hoogstwaarschijnlijk de noodzaak om protonen door het binnenste mitochondriale membraan te pompen in een poging om de ?? m te behouden, die cruciaal is voor de functionele integriteit van dit organel. De omkering van de functie van de F1F0-ATPase wordt ook bewezen door de waargenomen mitochondriale depolarisatie na toediening van oligomycine aan Nrf2-KO-cellen, wat in schril contrast staat met de hyperpolarisatie die optreedt in hun WT- of Keap1-deficiënte tegenhangers [35]. Over het algemeen lijkt het erop dat ATP onder omstandigheden van Nrf2-deficiëntie voornamelijk wordt geproduceerd in glycolyse, en deze ATP wordt vervolgens gedeeltelijk gebruikt door de F1F0-ATPase om de ?? m te behouden.
Nrf2 verbetert mitochondriale vetzuuroxidatie
Het effect van Nrf2-deficiëntie op de ?? m is vooral uitgesproken wanneer cellen worden geïncubeerd in medium zonder glucose, en de ?? m is ~ 50% lager in Nrf2-KO vergeleken met WT-cellen [35]. Onder omstandigheden van glucosetekort is mitochondriale vetzuuroxidatie (FAO) een belangrijke leverancier van substraten voor ademhaling en oxidatieve fosforylering, wat suggereert dat Nrf2 FAO kan beïnvloeden. Inderdaad, de efficiëntie van FAO voor zowel het lange-keten (C16: 0) verzadigde vetzuur palmitinezuur als het korte-keten (C6: 0) hexaanzuur is hoger in Keap1-KO MEF's en geïsoleerde hart- en lever mitochondriën dan in hun WT-tegenhangers, terwijl het lager is in Nrf2-KO-cellen en mitochondriën [65]. Deze effecten zijn ook zeer relevant voor mensen: inderdaad, metabolische veranderingen die duiden op een betere integratie van FAO met de activiteit van de TCA-cyclus zijn gemeld in menselijke interventiestudies met diëten die rijk zijn aan glucoraphanine, de voorloper van de klassieke Nrf2-activator sulforafaan [ 66].
Tijdens de eerste stap van mitochondriale FAO, verlaat de pro-R-waterstof van de β-koolstof zich als een hydride die de FAD-cofactor reduceert tot FADH2, dat op zijn beurt elektronen overbrengt naar ubiquinon (UbQ) in de ademhalingsketen, wat uiteindelijk bijdraagt aan de ATP-productie. . Terwijl stimulatie van FAO door palmitoylcarnitine in afwezigheid van glucose de verwachte toename van de ATP-niveaus in WT- en Keap1-KO-cellen veroorzaakt, waarbij de ATP-stijging sneller is in Keap1-KO-cellen, produceert dezelfde behandeling geen ATP-veranderingen in Nrf2-KO MEF's [65]. Dit experiment toont aan dat, in afwezigheid van Nrf2, FAO wordt onderdrukt, en bovendien impliceert het onderdrukking van FAO als een van de redenen voor de lagere ATP-niveaus onder omstandigheden van Nrf2-deficiëntie [35], [64].
Met name menselijke 293-T-cellen waarin Nrf2 tot zwijgen is gebracht hebben een lagere expressie van CPT1 en CPT2 [67], twee isovormen van carnitine palmitoyltransferase (CPT), het snelheidsbeperkende enzym in mitochondriale FAO. In overeenstemming zijn de mRNA-niveaus van Cpt1 lager in levers van Nrf2-KO in vergelijking met WT-muizen [68]. CPT katalyseert de overdracht van de acylgroep van een langketenige vetacyl-CoA van co-enzym A naar l-carnitine en maakt aldus de invoer van acylcarnitine uit het cytoplasma in de mitochondriën mogelijk. Hoewel dit tot op heden niet is onderzocht, is het mogelijk dat naast de transcriptie-effecten op CPT1-expressie, Nrf2 ook de functie van dit enzym kan beïnvloeden door de niveaus van de belangrijkste allostere remmer, malonyl-CoA, te regelen. Dit komt omdat, door een mechanisme dat momenteel onduidelijk is, negatieff2 de expressie van stearoyl CoA-desaturase (SCD) [69] en citraatlyase (CL) [69], [70] negatief reguleert. Merkwaardig genoeg leidt knock-out of remming van SCD tot verhoogde fosforylatie en activering van AMP-geactiveerde proteïnekinase (AMPK) [71], [72], [73], en er kan gespeculeerd worden dat, in de afwezigheid van Nrf2, de SCD-niveaus zal toenemen, waardoor de AMPK-activiteit wordt verlaagd. Dit zou verder kunnen worden verergerd door de verlaagde eiwitniveaus van AMPK die zijn waargenomen in levers van Nrf2-KO-muizen [68], een bevinding die nauw overeenkomt met de verhoogde AMPK-niveaus, die zijn gemeld in levers van Keap1-KD muizen [74]. Eén gevolg van de verlaagde AMPK-activiteit is de verlichting van de remmende fosforylering (aan Ser79) van acetyl-CoA-carboxylase (ACC) [75], die verder transcriptie opgereguleerd zou kunnen worden in de afwezigheid van Nrf2 omdat het wordt gedownreguleerd door Nrf2-activering [70 ]. De hoge ACC-activiteit, in combinatie met de upregulated CL-expressie die de productie van acetyl-CoA, het substraat voor ACC, zal verhogen, kan uiteindelijk de niveaus van het ACC-product, malonyl-CoA, verhogen. De hoge niveaus van malonyl-CoA remmen CPT, waardoor het transport van vetzuren naar de mitochondriën wordt verminderd. Ten slotte reguleert Nrf2 de expressie van CD36 [76], een translocase die vetzuren over het plasma en mitochondriale membranen importeert, positief. Eén mechanisme waarmee Nrf2 de efficiëntie van mitochondriaal FAO kan beïnvloeden, is dus door de invoer van langketenige vetzuren in de mitochondriën te reguleren.
Naast directe transcriptionele regulatie, kan Nrf2 ook de efficiëntie van mitochondriale FAO veranderen door zijn effecten op het cellulaire redoxmetabolisme. Dit kan met name relevant zijn wanneer de Nrf2-activiteit laag of afwezig is, waarbij de cellulaire redoxstatus verschuift naar de geoxideerde toestand. Inderdaad, verschillende FAO-enzymen zijn geïdentificeerd als gevoelig voor redoxveranderingen. Eén zo'n enzym is acyl-CoA-dehydrogenase met zeer lange ketens (VLCAD), dat meer dan 80% bijdraagt aan de palmitoyl-CoA-dehydrogeneringsactiviteit in menselijke weefsels [77]. Interessant is dat Hurd et al. [78] hebben aangetoond dat VLCAD cysteïne-residuen bevat die hun redox-toestand aanzienlijk veranderen na blootstelling van geïsoleerde mitochondria van het rattenhart aan H2O2. Bovendien verbetert S-nitrosylatie van muriene hepatische VLCAD bij Cys238 de katalytische efficiëntie van het enzym [79], en het is waarschijnlijk dat oxidatie van hetzelfde cysteïne het tegenovergestelde effect kan hebben, waardoor uiteindelijk de efficiëntie van mitochondriaal FAO wordt verlaagd. Het is daarom mogelijk dat, hoewel de expressieniveaus van VLCAD niet significant verschillen in WT, Nrf2-KO of Keap1-KO MEF's [65], de enzymactiviteit van VLCAD lager zou kunnen zijn in de afwezigheid van Nrf2 vanwege de hogere niveaus van ROS.
Op basis van al deze bevindingen kan worden voorgesteld dat (Fig. 3): bij afwezigheid van Nrf2 de NADPH-niveaus lager zijn als gevolg van verminderde expressie van ME1, IDH1, G6PD en PGD. De niveaus van gereduceerd glutathion zijn ook lager als gevolg van verminderde expressie van enzymen die deelnemen aan de biosynthese en regeneratie en de lagere niveaus van NADPH die nodig zijn voor de omzetting van het geoxideerde naar de gereduceerde vorm van glutathion. De lage expressie van ME1 zal de pool van pyruvaat die de mitochondriën binnendringt verminderen, waarbij glycolyse de belangrijkste bron van pyruvaat wordt. De generatie van NADH is langzamer, wat leidt tot verminderde activiteit van complex I en verhoogde mitochondriale ROS-productie. De reductie van FAD tot FADH2 is ook langzamer, althans gedeeltelijk als gevolg van minder efficiënte vetzuuroxidatie, waardoor de elektronenstroom van FADH2 naar UbQ en naar complex III in gevaar komt. Aangezien UbQH2 een activator is van succinaat dehydrogenase [80], kan het vertragen van de vorming ervan de enzymactiviteit van succinaat dehydrogenase verlagen. De verhoogde niveaus van superoxide en waterstofperoxide kunnen de activiteit van complex II verder remmen [81]. De lagere efficiëntie van vetzuuroxidatie draagt bij aan de verminderde beschikbaarheid van substraat voor mitochondriale ademhaling en ATP-productie bij oxidatieve fosforylering. Als compensatiemechanisme wordt de glycolyse verbeterd. ATP-synthase functioneert omgekeerd, als een ATPase, in een poging de ?? m te behouden.
Nrf2 en mitochondriale biogenese
Er is gerapporteerd dat, in vergelijking met WT, de levers van Nrf2-KO-muizen een lager mitochondriaal gehalte hebben (zoals bepaald door de verhouding tussen mitochondriaal en nucleair DNA); dit wordt verder verminderd met een 24-uur vasten in zowel WT- als Nrf2-KO-muizen; daarentegen, hoewel niet verschillend van WT onder normale voedingsomstandigheden, wordt het mitochondriale gehalte in muizen met hoge Nrf2-activiteit niet beïnvloed door vasten [82]. Interessant is dat suppletie met de Nrf2-activator (R) -? - liponzuur [83], [84], [85] de mitochondriale biogenese in 3T3-L1-adipocyten bevordert [86]. Twee klassen van nucleaire transcriptionele regulatoren spelen een cruciale rol in de mitochondriale biogenese. De eerste klasse zijn transcriptiefactoren, zoals nucleaire ademhalingsfactoren11 en 2, die de expressie regelen van genen die coderen voor subeenheden van de vijf ademhalingscomplexen, mitochondriale translatiecomponenten en biosynthetische heem-enzymen die zijn gelokaliseerd in de mitochondriale matrix [88]. Piantadosi et al. [89] hebben aangetoond dat de Nrf2-afhankelijke transcriptionele opregulatie van nucleaire respiratoire factor 1 mitochondriale biogenese bevordert en beschermt tegen de cytotoxiciteit van het cardiotoxische anthracycline chemotherapeutische middel doxorubicine. Daarentegen Zhang et al. [82] hebben gerapporteerd dat genetische activering van Nrf2 geen invloed heeft op de basale mRNA-expressie van nucleaire respiratoire factor 1 in de muizenlever.
De tweede klasse van nucleaire transcriptionele regulatoren met kritische functies in mitochondriale biogenese zijn transcriptionele co-activatoren, zoals peroxisoomproliferator-geactiveerde receptor? coactivators (PGC) 1? en 1 ?, die een interactie aangaan met transcriptiefactoren, de basale transcriptionele en RNA-splitsingsmachines, en histon-modificerende enzymen [88], [90], [91]. De expressie van de PGC1-familie van co-activatoren wordt beïnvloed door talrijke omgevingssignalen. Behandeling van menselijke fibroblasten met de Nrf2-activator sulforafaan veroorzaakt een toename van de mitochondriale massa en inductie van PGC1? en PGC1? [92], hoewel de mogelijke afhankelijkheid van Nrf2 in deze studie niet werd onderzocht. Diabetische muizen waarbij Nrf2 wordt geactiveerd door Keap1-gen hypomorfe knockdown (db / db: Keap1flox / ?: Nrf2 + / +) of verstoord (db / db: Keap1flox / ?: Nrf2? /?), Hebben een lagere hepatische PGC1? expressieniveaus dan controledieren (db / db: Keap1flox / +: Nrf2 + / +) [93]. Geen verschillen in de mRNA-niveaus voor PGC1? worden gezien in levers van niet-diabetische muizen die ofwel WT of Nrf2-KO zijn, terwijl deze niveaus lager zijn bij dieren die Nrf2 tot overexpressie brengen (Keap1-KD en leverspecifieke Keap1-KO) [82]. Met name, een 24-uurs vasten verhoogt de niveaus van PGC1? mRNA in de levers van muizen van alle genotypen, maar de toename is significant groter in levers van Nrf2-KO vergeleken met muizen die WT of Nrf2 tot overexpressie brengen. In vergelijking met WT vertonen Nrf2-KO-muizen die septische infectie of acuut longletsel als gevolg van infectie ervaren een verzwakte transcriptionele opregulatie van nucleaire respiratoire factor 1 en PGC1? [94], [95]. Samen suggereren deze waarnemingen dat de rol van Nrf2 bij het handhaven van de niveaus van zowel nucleaire respiratoire factor 1 als PGC1? is complex en wordt het meest prominent onder stressomstandigheden.
Naast de expressie van genen die coderen voor mitochondriale eiwitten, vereist mitochondriale biogenese de synthese van nucleotiden. Genetische activering van Nrf2 verbetert de biosynthese van purine door de pentosefosfaatroute en het metabolisme van folaat en glutamine te verhogen, vooral in snel prolifererende cellen (Fig. 2) [24]. Analyse van het transcriptoom van mutant Drosophila deficiënt voor het mitochondriale serine / threonine proteïne kinase PTEN-geïnduceerde vermeende kinase 1 (PINK1) heeft aangetoond dat mitochondriale disfunctie leidt tot de transcriptionele opregulatie van genen die het nucleotide metabolisme beïnvloeden [96], wat suggereert dat de verbeterde nucleotide biosynthese vertegenwoordigt een mechanisme voor bescherming tegen de neurotoxische gevolgen van PINK1-deficiëntie. Nrf2 reguleert de expressie van fosforibosylpyrofosfaatamidotransferase (PPAT), dat de toegang tot de de novo purine nucleotide biosynthetische route katalyseert, en mitochondriaal methyleentetrahydrofolaat dehydrogenase 2 (MTHFD2) (figuur 2). Dit laatste is een bifunctioneel enzym met dehydrogenase- en cyclohydrolase-activiteiten dat cruciaal is voor het leveren van zowel glycine als formiaat als bronnen van één-koolstofeenheden voor purinebiosynthese in snelgroeiende cellen [97]. Het is daarom waarschijnlijk dat Nrf2-activering beschermend zou kunnen zijn en mitochondriale disfunctie bij PINK1-deficiëntie zou kunnen omkeren. Inderdaad, farmacologische activering van Nrf2 door sulforafaan, of de triterpenoïde RTA-408, herstelt en beschermt PINK1-deficiënte cellen tegen dopamine-toxiciteit [98]. Hoewel de onderliggende mechanismen complex lijken, geven deze bevindingen samen aan dat Nrf2-activiteit de mitochondriale biogenese kan beïnvloeden door de expressieniveaus van kritische transcriptiefactoren en co-activatoren te beïnvloeden, evenals door de nucleotidebiosynthese te verbeteren.
Nrf2 en mitochondriale integriteit
Hoewel er niet altijd direct bewijsmateriaal beschikbaar is, zijn er sterke aanwijzingen dat Nrf2 belangrijk is voor de mitochondriale integriteit, vooral onder omstandigheden van oxidatieve stress. Mitochondria geïsoleerd uit de hersenen en lever van ratten waaraan een enkele dosis van de Nrf2-activator sulforafaan was toegediend, zijn resistent tegen het openen van de mitochondriale permeabiliteitsovergangsporie (mPTP) veroorzaakt door het oxidatiemiddel tert-butylhydroperoxide [99], [100]. Het mPTP, een complex dat het mogelijk maakt dat het mitochondriale binnenmembraan permeabel wordt voor moleculen met massa tot 1500 Da, werd recent geïdentificeerd als gevormd uit dimeren van het F0F1-ATP-synthase [101]. De door sulforafaan gemedieerde resistentie tegen mPTP-opening correleert met toegenomen antioxidantafweer en de niveaus van mitochondriaal GSH, glutathionperoxidase 1, malisch enzym 3 en thioredoxine 2 zijn alle opgereguleerd in mitochondriale fracties geïsoleerd uit met sulforaphane behandelde dieren [100].
Mitochondriale proteïneschade en verminderde ademhaling veroorzaakt door het elektrofiele lipideperoxidatieproduct 4-hydroxy-2-nonenal worden verzwakt in mitochondriën geïsoleerd uit de hersenschors van met sulforafaan behandelde muizen [102]. In renale epitheelcellen van de rat en in de nieren beschermt sulforafaan tegen cisplatine en gentamicine-geïnduceerde toxiciteit en verlies van ?? m [103], [104]. Bescherming tegen een reeks oxidanten (superoxide, waterstofperoxide, peroxynitriet) en elektrofielen (4-hydroxy-2-nonenal en acroleïne) en een toename van de mitochondriale antioxidantafweer zijn ook waargenomen bij behandeling van gladde spiercellen van de aorta van de rat met sulforafaan [105 ]. In een model van door contrast geïnduceerd acuut nierletsel, werd onlangs aangetoond dat ischemische preconditionering van de ledematen beschermende effecten heeft, waaronder remming van de opening van de mPTP en mitochondriale zwelling, door activering van Nrf2 als gevolg van de remming van GSK3? [106].
Mitofagie, het proces waarbij disfunctionele mitochondriën selectief worden verzwolgen door autofagosomen en afgegeven aan lysosomen om te worden afgebroken en gerecycled door de cel, is essentieel voor mitochondriale homeostase [107], [108]. Hoewel er geen oorzakelijk verband tussen Nrf2 en mitofagie is vastgesteld, zijn er aanwijzingen dat de transcriptiefactor belangrijk kan zijn bij mitochondriale kwaliteitscontrole door een rol te spelen bij mitofagie. Dit kan vooral prominent zijn onder omstandigheden van oxidatieve stress. Dus, in een model van sepsis, worden de toenames in de niveaus van de autofagosoommarker MAP1 lichte keten 3-II (LC3-II) en het ladingeiwit p62 24 uur na infectie onderdrukt in Nrf2-KO vergeleken met WT-muizen [109] . Een kleine molecuul inductor van mitofagie (p62-gemedieerde mitofagie inductor genoemd, PMI) werd onlangs ontdekt; deze 1,4-difenyl-1,2,3-triazoolverbinding was oorspronkelijk ontworpen als een Nrf2-activator die de interactie van de transcriptiefactor met Keap1 verstoort [110]. Net als cellen waarin Nrf2 genetisch opgereguleerd is (Keap1-KD of Keap1-KO), hebben cellen die zijn blootgesteld aan PMI een hogere rustwaarde. Belangrijk is dat de toename in mitochondriale LC3-lokalisatie die wordt waargenomen na PMI-behandeling van WT-cellen niet optreedt in Nrf2-KO-cellen, wat de betrokkenheid van Nrf2 suggereert.
Tenslotte heeft ultrastructurele analyse van levercoupes de aanwezigheid van gezwollen mitochondria met gereduceerde crista en verstoorde membranen in hepatocyten van Nrf2-KO, maar niet WT, muizen die gedurende 24 weken een vetrijk dieet hadden gekregen, onthuld; met name deze levers vertonen duidelijk bewijs van oxidatieve stress en ontsteking [68]. Er kan worden geconcludeerd dat Nrf2 een cruciale rol speelt bij het handhaven van de mitochondriale integriteit onder omstandigheden van oxidatieve en inflammatoire stress.
Sulforafaan en de effecten ervan op kanker, mortaliteit, veroudering, hersenen en gedrag, hartaandoeningen en meer
Isothiocyanaten zijn enkele van de belangrijkste plantaardige stoffen die u in uw dieet kunt krijgen. In deze video maak ik de meest uitgebreide case voor hen die ooit is gemaakt. Korte aandachtsspanne? Ga naar uw favoriete onderwerp door op een van de onderstaande tijdspunten te klikken. Volledige tijdlijn hieronder.
Belangrijkste secties:
00: 01: 14 - Kanker en sterfte
00: 19: 04 - Veroudering
00: 26: 30 - Hersenen en gedrag
00: 38: 06 - Laatste overzicht
00: 40: 27 - Dosis
Volledige tijdlijn:
00: 00: 34 - Introductie van sulforafaan, een belangrijk aandachtspunt van de video.
00: 01: 14 - Kruisbloemige groenteconsumptie en verlagingen van mortaliteit door alle oorzaken.
00: 02: 12 - Risico op prostaatkanker.
00: 02: 23 - Blaaskankerrisico.
00: 02: 34 - Longkanker bij rokers risico.
00: 02: 48 - Risico op borstkanker.
00: 03: 13 - Hypothetisch: wat als je al kanker hebt? (Interventie)
00: 03: 35 - Aannemelijk mechanisme dat de associatieve gegevens over kanker en mortaliteit stuurt.
00: 04: 38 - Sulforafaan en kanker.
00: 05: 32 - Dierlijk bewijs dat een sterk effect van extract van broccolispruit toont op de ontwikkeling van blaastumoren bij ratten.
00: 06: 06 - Effect van directe suppletie van sulforafaan bij prostaatkankerpatiënten.
00: 07: 09 - Bioaccumulatie van isothiocyanaatmetabolieten in daadwerkelijk borstweefsel.
00: 08: 32 - Remming van stamcellen van borstkanker.
00: 08: 53 - Geschiedenisles: brassica's werden vastgesteld met gezondheidseigenschappen zelfs in het oude Rome.
00: 09: 16 - het vermogen van Sulforaphane om de kankerverwekkende uitscheiding te verbeteren (benzeen, acroleïne).
00: 09: 51 - NRF2 als een genetische switch via antioxidantresponselementen.
00: 10: 10 - Hoe NRF2-activering de carcinogene excretie via glutathion-S-conjugaten verbetert.
00: 10: 34 - Spruitjes verhogen glutathion-S-transferase en verminderen DNA-schade.
00: 11: 20 - Broccolispruitendrank verhoogt de benzeenuitscheiding met 61%.
00: 13: 31 - Broccoli-spruithomogenaat verhoogt antioxiderende enzymen in de bovenste luchtwegen.
00: 15: 45 - Kruisbloemige groenteconsumptie en sterfte aan hart- en vaatziekten.
00: 16: 55 - Broccolispruitpoeder verbetert de bloedlipiden en het algemene risico op hartziekten bij type 2 diabetici.
00: 19: 04 - Begin van het verouderingsgedeelte.
00: 19: 21 - Met Sulforaphane verrijkte voeding verlengt de levensduur van kevers van 15 tot 30% (onder bepaalde omstandigheden).
00: 20: 34 - Belang van lage ontsteking voor een lang leven.
00: 22: 05 - Kruisbloemige groenten en broccoli-kiempoeder lijken een breed scala aan ontstekingsmarkers bij mensen te verminderen.
00: 24: 14 - Muisstudies suggereren dat sulforaphane de adaptieve immuunfunctie op oudere leeftijd zou kunnen verbeteren.
00: 25: 18 - Sulforafaan verbeterde de haargroei in een muismodel van kalend. Afbeelding bij 00: 26: 10.
00: 26: 30 - Begin van de sectie hersenen en gedrag.
00: 27: 18 - Effect van extract van broccolispruiten op autisme.
00: 27: 48 - Effect van glucoraphanine op schizofrenie.
00: 28: 17 - Begin van depressie discussie (plausibel mechanisme en studies).
00: 31: 21 - Muisstudie met behulp van 10 verschillende modellen van stress-geïnduceerde depressie laten sulforafaan zien, even effectief als fluoxetine (prozac).
00: 32: 00 - Onderzoek toont aan dat directe inname van glucoraphanine bij muizen even effectief is bij het voorkomen van depressies als gevolg van een sociaal nederlaagstressmodel.
00: 33: 01 - Begin van de sectie neurodegeneratie.
00: 33: 30 - Sulforafane en de ziekte van Alzheimer.
00: 33: 44 - Sulforafane en de ziekte van Parkinson.
00: 33: 51 - De ziekte van Sulforaphane en Hungtington.
00: 34: 43 - Begin van een traumatisch gedeelte over hersenletsel.
00: 35: 01 - Sulforafaan wordt geïnjecteerd onmiddellijk nadat TBI het geheugen verbetert (muisstudie).
00: 35: 55 - Sulforafaan en neuronale plasticiteit.
00: 36: 32 - Sulforafaan verbetert het leren in het model van type II diabetes bij muizen.
00: 37: 19 - Sulforafaan en duchenne spierdystrofie.
00: 37: 44 - Myostatinremming in spieratlanticellen (in vitro).
00: 38: 06 - Late video recapitulatie: sterfte en kanker, DNA-schade, oxidatieve stress en ontsteking, uitscheiding van benzeen, cardiovasculaire aandoeningen, type II diabetes, effecten op de hersenen (depressie, autisme, schizofrenie, neurodegeneratie), NRF2-route.
00: 40: 27 - Gedachten over het uitzoeken van een dosis broccoli of sulforafaan.
00: 41: 01 - Anekdotes over ontspruiten thuis.
00: 43: 14 - Op kooktemperaturen en sulforafaanactiviteit.
00: 43: 45 - Darmbacterieomzetting van sulforafaan uit glucoraphanine.
00: 44: 24 - Supplementen werken beter in combinatie met actieve myrosinase van groenten.
00: 44: 56 - Koken technieken en kruisbloemige groenten.
00: 46: 06 - Isothiocyanaten als goitrogens.
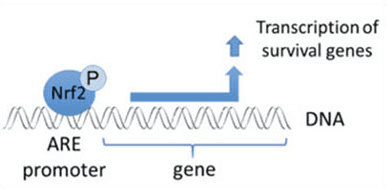
Nrf2 is een transcriptiefactor die een belangrijke rol speelt in het cellulaire antioxidante afweersysteem van het menselijk lichaam. Het op antioxidanten reagerende element, of ARE, is een regulerend mechanisme van genen. Veel onderzoeksstudies hebben aangetoond dat Nrf2, of NF-E2-gerelateerde factor 2, een breed scala van door ARE gestuurde genen in verschillende soorten cellen reguleert. Nrf2 bleek ook een essentiële rol te spelen bij cellulaire bescherming en anti-carcinogeniciteit, wat aantoont dat Nrf2 een effectieve behandeling kan zijn bij de behandeling van neurodegeneratieve ziekten en kankers die vermoedelijk door oxidatieve stress worden veroorzaakt. Dr. Alex Jimenez DC, CCST Insight
Slotopmerkingen
Hoewel er nog veel vragen open blijven, geeft het beschikbare experimentele bewijs duidelijk aan dat Nrf2 een belangrijke speler is in het behoud van mitochondriale homeostase en structurele integriteit. Deze rol wordt vooral kritisch onder omstandigheden van oxidatieve, elektrofiele en ontstekingsstress wanneer het vermogen om door Nrf2 gemedieerde cytoprotectieve responsen op te winden, de algehele gezondheid en overleving van de cel en het organisme beïnvloedt. De rol van Nrf2 in de mitochondriale functie vertegenwoordigt een andere laag van de brede cytoprotectieve mechanismen georkestreerd door deze transcriptiefactor. Aangezien veel pathologische aandoeningen bij mensen oxidatieve stress, ontsteking en mitochondriale disfunctie hebben als essentiële componenten van hun pathogenese, houdt farmacologische activering van Nrf2 veelbelovend in voor ziektepreventie en behandeling. Alomvattend begrip van de precieze mechanismen waarmee Nrf2 de mitochondriale functie beïnvloedt, is essentieel voor een rationeel ontwerp van toekomstige klinische onderzoeken en kan nieuwe biomarkers bieden voor het bewaken van de therapeutische werkzaamheid.
Het doel van het bovenstaande artikel was om de opkomende rol van Nrf2 in de mitochondriale functie te bespreken en aan te tonen. Nrf2 of kernfactor erytroïde 2-gerelateerde factor, is een opkomende regulator van cellulaire resistentie tegen oxidanten die kan bijdragen aan oxidatieve stress, de cellulaire functie kan aantasten en kan leiden tot de ontwikkeling van toxiciteit, chronische ziekten en zelfs kanker. Hoewel de productie van oxidanten in het menselijk lichaam verschillende doelen kan dienen, waaronder celdeling, ontsteking, immuunfunctie, autofagie en stressreactie, is het essentieel om hun overproductie onder controle te houden om gezondheidsproblemen te voorkomen. De reikwijdte van onze informatie is beperkt tot chiropractie en problemen met de wervelkolom. Om het onderwerp te bespreken, kunt u het aan Dr. Jimenez vragen of contact met ons opnemen via 915-850-0900 .
Rugpijn is een van de meest voorkomende oorzaken van arbeidsongeschiktheid en gemiste werkdagen wereldwijd. Rugpijn is de tweede meest voorkomende reden voor doktersbezoeken, alleen overtroffen door bovenste luchtweginfecties. Ongeveer 80 procent van de bevolking zal gedurende zijn leven minstens één keer rugpijn ervaren. De wervelkolom is een complexe structuur die bestaat uit botten, gewrichten, ligamenten en spieren, naast andere zachte weefsels. Hierdoor kunnen verwondingen en / of verergerde omstandigheden, zoals herniated discs, kan uiteindelijk leiden tot symptomen van rugpijn. Sportblessures of auto-ongelukken zijn vaak de meest voorkomende oorzaak van rugpijn, maar soms kunnen de eenvoudigste bewegingen pijnlijke gevolgen hebben. Gelukkig kunnen alternatieve behandelingsopties, zoals chiropractische zorg, rugpijn helpen verlichten door het gebruik van spinale aanpassingen en handmatige manipulaties, waardoor uiteindelijk de pijnverlichting wordt verbeterd. �
Nrf2 ondersteunt de activering van een groep antioxidanten en ontgiftende enzymen en genen die het menselijk lichaam beschermen tegen de gevolgen van gezondheidsproblemen die gepaard gaan met verhoogde niveaus van oxidatieve stress, zoals de ziekte van Alzheimer. Van een verscheidenheid aan natuurlijke stoffen is aangetoond dat ze de Nrf2-route activeren, wat kan helpen de symptomen van neurodegeneratieve ziekten te beheersen. Het doel van het onderstaande artikel is om de cruciale rol van Nrf2, veroorzaakt door chronische ontsteking, te bespreken.
Abstract
Ontsteking is het meest voorkomende kenmerk van veel chronische ziekten en complicaties, terwijl het een cruciale rol speelt bij carcinogenese. Verschillende onderzoeken hebben aangetoond dat Nrf2 bijdraagt aan het ontstekingsremmende proces door de rekrutering van ontstekingscellen te orkestreren en de genexpressie te reguleren via het antioxidant respons element (ARE). De Keap1 (Kelch-achtige ECH-geassocieerd eiwit) / Nrf2 (NF-E2 p45-gerelateerde factor 2) / ARE-signaleringsroute reguleert voornamelijk ontstekingsremmende genexpressie en remt de progressie van ontsteking. Daarom is de identificatie van nieuwe Nrf2-afhankelijke ontstekingsremmende fytochemicaliën een belangrijk punt geworden bij het ontdekken van geneesmiddelen. In deze review bespreken we de leden van de Keap1 / Nrf2 / ARE-signaalroute en zijn stroomafwaartse genen, de effecten van deze route op diermodellen van ontstekingsziekten en de overspraak met de NF-? B-route. Daarnaast bespreken we ook de regulatie van NLRP3-inflammasoom door Nrf2. Daarnaast vatten we het huidige scenario samen van de ontwikkeling van ontstekingsremmende fytochemicaliën en andere die de Nrf2 / ARE-signaleringsroute bemiddelen.
Ontsteking is een complex proces dat optreedt wanneer weefsels worden geïnfecteerd of gewond door schadelijke stimuli, zoals ziekteverwekkers, schade of irriterende stoffen. Immuuncellen, bloedvaten en moleculaire bemiddelaars zijn betrokken bij deze beschermende respons [1]. Ontsteking is ook een pathologisch fenomeen dat geassocieerd is met een verscheidenheid van ziektetoestanden die voornamelijk worden veroorzaakt door fysische, chemische, biologische en psychologische factoren. Het doel van ontsteking is het beperken en elimineren van de oorzaken van cellulaire schade, het verwijderen en / of absorberen van necrotische cellen en weefsels en het starten van weefselherstel. Er worden twee verschillende vormen van ontsteking onderscheiden: acuut en chronisch. Acute ontsteking is zelflimiterend en gunstig voor de gastheer, maar langdurige chronische ontsteking is een veel voorkomend kenmerk van veel chronische ziekten en complicaties. Directe infiltratie door vele mononucleaire immuuncellen zoals monocyten, macrofagen, lymfocyten en plasmacellen, evenals de productie van inflammatoire cytokinen, leiden tot chronische ontsteking. Erkend wordt dat chronische ontsteking een cruciale rol speelt in carcinogenese [2]. In het algemeen werken zowel pro- als ontstekingsremmende signaleringsroutes samen in het normale ontstekingsproces.
Bij het pathologische ontstekingsproces worden eerst mestcellen, monocyten, macrofagen, lymfocyten en andere immuuncellen geactiveerd. Vervolgens worden de cellen gerekruteerd op de plaats van verwonding, wat resulteert in het genereren van reactieve zuurstofsoorten (ROS) die macromoleculen, waaronder DNA, beschadigen. Tegelijkertijd produceren deze ontstekingscellen ook grote hoeveelheden ontstekingsmediatoren zoals cytokinen, chemokinen en prostaglandinen. Deze mediatoren rekruteren macrofagen verder naar gelokaliseerde ontstekingsplaatsen en activeren direct meerdere signaaltransductiecascades en transcriptiefactoren die met ontsteking zijn geassocieerd. De NF-? B (nucleaire factor kappa B), MAPK (mitogeen-geactiveerde proteïne kinase) en JAK (janus kinase) -STAT (signaaltransducers en activatoren van transcriptie) signaleringsroutes zijn betrokken bij de ontwikkeling van de klassieke ontstekingsroute. [3], [4], [5]. Eerdere studies hebben aangetoond dat de transcriptiefactor Nrf2 (NF-E2 p45-gerelateerde factor 2) de expressie reguleert van fase II-ontgiftende enzymen, waaronder NADPH, NAD (P) H-chinonoxidoreductase 1, glutathionperoxidase, ferritine, heem-oxygenase-1 (HO -1), en antioxidantgenen die cellen beschermen tegen verschillende verwondingen via hun ontstekingsremmende effecten, en zo het verloop van de ziekte beïnvloeden [6], [7], [8].
Gezien deze opmerkelijke bevindingen heeft de ontwikkeling van gerichte therapeutische geneesmiddelen voor ontstekingsziekten via signaleringsroutes de laatste jaren veel belangstelling getrokken. In deze review vatten we onderzoek samen naar de Keap1 (Kelch-achtige ECH-geassocieerd eiwit) / Nrf2 (NF-E2 p45-gerelateerde factor 2) / ARE (antioxidant responselement) signaleringsroute bij ontstekingen.
Structuur en regulatie van Nrf2
Keap1-afhankelijke Nrf2-verordening
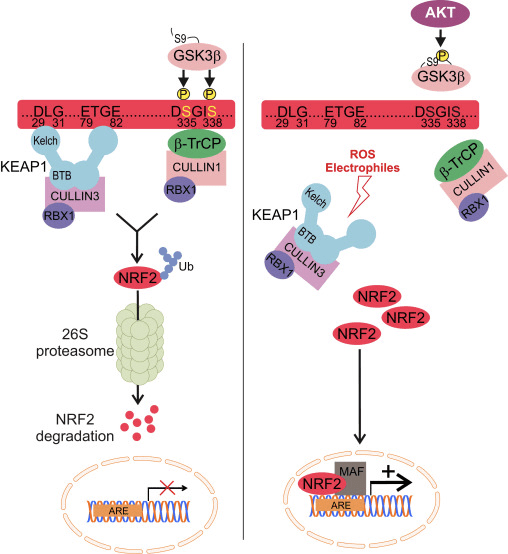
Nrf2 behoort tot de Cap n Collar (CNC) -subfamilie en omvat in zeven functionele domeinen Neh (Nrf2-ECH-homologie) 1 tot Neh7 [9], [10]. Neh1 is een CNC-bZIP-domein waarmee Nrf2 kan heterodimeriseren met klein musculoaponeurotisch fibrosarcoom (Maf) -eiwit, DNA en andere transcriptiepartners, en ook een nucleair complex kan vormen met het ubiquitine-conjugerende enzym UbcM2 [11], [12]. Neh2 bevat twee belangrijke motieven die bekend staan als DLG en ETGE, die essentieel zijn voor de interactie tussen Nrf2 en zijn negatieve regulator Keap1 [13], [14].
Keap1 is een substraatadapter voor op cullin gebaseerde E3 ubiquitine-ligase, die de transcriptieactiviteit van Nrf2 remt via ubiquitinatie en proteasomale afbraak onder normale omstandigheden [15], [16], [17]. De KELCH-domeinen van het Keap1-homodimeer binden met de DLG- en ETGE-motieven van het Nrf2-Neh2-domein in het cytosol, waar ETGE werkt als een scharnier met hogere affiniteit en DLG fungeert als een grendel [18]. Onder oxidatieve stress of bij blootstelling aan Nrf2-activatoren dissocieert Nrf2 van Keap1-binding vanwege de thiol-modificatie van Keap1-cysteïne-residuen die uiteindelijk Nrf2-ubiquitinatie en proteasomale afbraak [19] voorkomt. Vervolgens verplaatst Nrf2 zich naar de kern, heterodimeriseert met kleine Maf-eiwitten en transactiveert een ARE-batterij van genen (Fig. 1A). Het carboxy-uiteinde van Neh3 fungeert als een transactivatiedomein door interactie met de transcriptie-co-activator bekend als CHD6 (chromo-ATPase / helicase-DNA-bindend eiwit) [20]. Neh4 en Neh5 werken ook als transactivatiedomeinen, maar binden aan een andere transcriptionele co-activator bekend als CBP (cAMP-response-element-binding protein-binding protein) [21]. Bovendien werken Neh4 en Neh5 samen met de nucleaire cofactor RAC3 / AIB1 / SRC-3, wat leidt tot verbeterde op Nrf2 gerichte ARE-genexpressie [22]. Neh5 heeft een redox-gevoelig kern-export signaal dat cruciaal is voor de regulatie en cellulaire lokalisatie van Nrf2 [23].
Figuur 1 Keap1-afhankelijke en -onafhankelijke regeling van Nrf2. (A) Onder basale omstandigheden wordt Nrf2 gesekwestreerd met Keap1 door zijn twee motieven (ETGE en DLG) die leiden tot CUL3-gemedieerde ubiquitinatie gevolgd door proteasoomafbraak. Onder oxidatieve stress dissocieert Nrf2 van Keap1, verplaatst het zich naar de kern en activeert het de ARE-genbatterij. (B) GSK3 fosforyleert Nrf2 en dit vergemakkelijkt de herkenning van Nrf2 door? -TrCP voor CUL1-gemedieerde ubiquitinatie en daaropvolgende proteasoomafbraak. (C) p62 wordt afgezonderd met Keap1, wat leidt tot zijn autofagische afbraak, de bevrijding van Nrf2 en verhoogde Nrf2-signalering.
Keap1-onafhankelijke Nrf2-verordening
Opkomend bewijs heeft een nieuw mechanisme van Nrf2-regulering onthuld dat onafhankelijk is van Keap1. Het serinerijke Neh6-domein van Nrf2 speelt een cruciale rol in deze regulatie door met zijn twee motieven (DSGIS en DSAPGS) te binden aan? -Transducine-herhalend eiwit (? -TrCP) [24]. ? -TrCP is een substraatreceptor voor het Skp1 Cul1 Rbx1 / Roc1 ubiquitine-ligasecomplex dat zich richt op Nrf2 voor ubiquitinatie en proteasomale afbraak. Glycogeensynthase kinase-3 is een cruciaal eiwit dat betrokken is bij Keap1-onafhankelijke Nrf2-stabilisatie en -regulatie; het fosforyleert Nrf2 in het Neh6-domein om de herkenning van Nrf2 door p-TrCP en daaropvolgende eiwitafbraak te vergemakkelijken [25] (Fig. 1B).
Andere Nrf2-regelaars
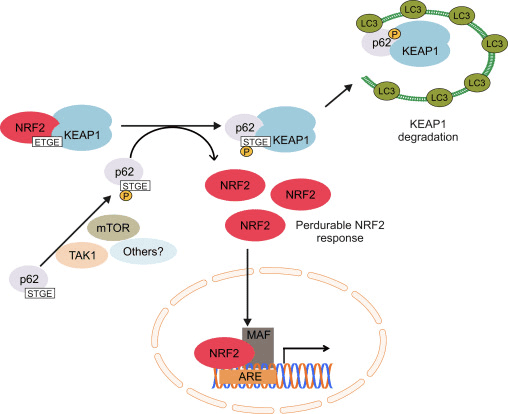
Een andere bewijslijn heeft een niet-canonieke route van p62-afhankelijke Nrf2-activering aangetoond waarbij p62 sequenties Keap1 sequestreert voor autofagische afbraak die uiteindelijk leidt tot de stabilisatie van Nrf2 en de transactivatie van Nrf2-afhankelijke genen [26], [27], [ 28], [29] (fig. 1C).
Accumulerend bewijs suggereert dat verschillende miRNA's een belangrijke rol spelen in de regulatie van de Nrf2-activiteit [30]. Sangokoya et al. [31] toonde aan dat miR-144 de Nrf2-activiteit direct downreguleert in de lymfoblast K562-cellijn, primaire menselijke erytroïde voorlopercellen en sikkelcelziekte-reticulocyten. Een andere interessante studie in menselijke borstepitheelcellen toonde aan dat miR-28 Nrf2 remt via een Keap1-onafhankelijk mechanisme [32]. Evenzo downreguleren miRNA's zoals miR-153, miR-27a, miR-142-5p en miR144 Nrf2-expressie in de neuronale SH-SY5Y-cellijn [33]. Singh et al. [34] toonde aan dat de ectopische expressie van miR-93 de expressie van Nrf2-gereguleerde genen vermindert in een 17? -Estradiol (E2) -geïnduceerd ratmodel van borstcarcinogenese.
Een recente ontdekking uit ons laboratorium identificeerde een endogene remmer van Nrf2 bekend als retinoïsche X-receptor alfa (RXR?). RXR? is een nucleaire receptor, interageert met het Neh7-domein van Nrf2 (aminozuurresiduen 209-316) via zijn DNA-bindende domein (DBD) en remt specifiek Nrf2-activiteit in de kern. Bovendien is gerapporteerd dat andere nucleaire receptoren, zoals peroxisoomproliferator-geactiveerde receptor-?, ER ?, oestrogeen-gerelateerde receptor-?, En glucocorticoïde receptoren ook endogene remmers zijn van Nrf2-activiteit [9], [10].
Ontstekingsremmende rol van de Nrf2 / HO-1-as
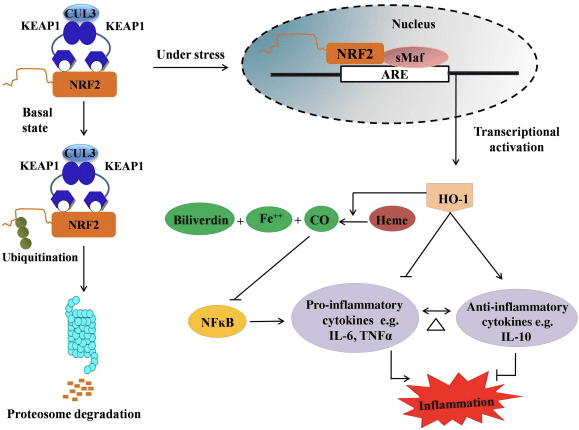
HO-1 is het induceerbare isovorm en snelheidsbeperkende enzym dat de afbraak van heem in koolmonoxide (CO) en vrij ijzer en biliverdine tot bilirubine katalyseert. Enzymatische afbraak van pro-inflammatoire, vrije heem, evenals de productie van ontstekingsremmende verbindingen zoals CO en bilirubine spelen een belangrijke rol bij het handhaven van de beschermende effecten van HO-1 (Fig. 2).
Figuur 2 Overzicht van de Nrf2 / HO-1-route. Onder basale omstandigheden bindt Nrf2 zich aan zijn repressor Keap1, wat leidt tot ubiquitinatie gevolgd door proteasoomafbraak. Tijdens oxidatieve stress transloceert vrij Nrf2 naar de kern, waar het dimeriseert met leden van de kleine Maf-familie en zich bindt aan ARE-genen zoals HO-1. Upregulated HO-1 katalyseert het heem in CO, bilirubine en vrij ijzer. CO werkt als een remmer van de NF-? B-route die leidt tot een verminderde expressie van pro-inflammatoire cytokines, terwijl bilirubine ook als antioxidant werkt. Bovendien remt HO-1 direct de pro-inflammatoire cytokines en activeert het de ontstekingsremmende cytokines, waardoor het ontstekingsproces in evenwicht wordt gebracht.
Nrf2 induceert het HO-1-gen door de mRNA- en eiwitexpressie te verhogen en het is een van de klassieke Nrf2-gereguleerde genen die veel wordt gebruikt in tal van in vitro en in vivo onderzoeken. Verschillende onderzoeken hebben aangetoond dat HO-1 en zijn metabolieten significante ontstekingsremmende effecten hebben die worden gemedieerd door Nrf2. Verhoging van HO-1-expressie die wordt gemedieerd door geactiveerde Nrf2 leidt tot de remming van NF? B-signalering resulteert in de verminderde intestinale mucosale schade en tight-junction-disfunctie in het mannelijke Sprague-Dawley-levertransplantatiemodel bij ratten [35]. Opwaartse regulatie van Nrf2-afhankelijke HO-1-expressie kan van muis afgeleide C2C12-myoblasten beschermen tegen H2O2-cytotoxiciteit [36]. Nrf2-afhankelijke HO-1 heeft een impact op door lipopolysaccharide (LPS) gemedieerde ontstekingsreacties in RAW264.7- of muis peritoneale macrofaag-afgeleide schuimcelmacrofagen. Nrf2-activiteit maakte het fenotype van schuimcelmacrofagen ongevoelig en voorkomt onmatige ontsteking van macrofagen, die een belangrijke rol spelen bij de progressie van atherosclerose [37]. De Nrf2 / HO-1-as beïnvloedt LPS-geïnduceerde muis BV2-microgliale cellen en muis hippocampale HT22-cellen, met impact op neuro-inflammatie. Upregulatie van HO-1 expressie via Nrf2 pathway in muis BV2 microgliacellen die celdood van muis hippocampale HT22 cellen verdedigen [38]. Bovendien verhogen op kobalt gebaseerde hybride moleculen (HYCO's) die een Nrf2-inductor combineren met een afgifte van koolmonoxide (CO) de Nrf2 / HO-1-expressie, maken CO vrij en oefenen ze ontstekingsremmende activiteit uit in vitro. HYCO's reguleren ook weefsel HO-1 en leveren CO in het bloed na toediening in vivo, wat hun potentiële gebruik tegen inflammatoire aandoeningen ondersteunt [39]. De opregulatie van Nrf2 / HO-1 vermindert de ontsteking door de efferocytische activiteit van macrofagen van muizen die zijn behandeld met taurinechlooramines te verhogen [40]. Al met al toonden de hierboven uiteengezette experimentele modellen aan dat de Nrf2 / HO-1-as een belangrijke rol speelt bij de ontstekingsremmende functie, wat suggereert dat Nrf2 een therapeutisch doelwit is bij met ontsteking geassocieerde ziekten.
Bovendien werken de bijproducten van HO-1, zoals CO, bilirubine, als een krachtige antioxidant tijdens oxidatieve stress en celbeschadiging [41], [42]; het onderdrukt auto-immune encefalomyelitis en hepatitis [43], [44]; en het beschermt muizen en ratten tegen endotoxische shock door het genereren van iNOS en NO [45], [46], [47] te voorkomen. Bovendien vermindert Bilirubine de activering en het functioneren van het endotheel [48]. Interessant is dat bilirubine de transmigratie van endotheliale leukocyten vermindert via adhesiemolecuul-1 [49]. Deze specifieke referenties die niet alleen HO-1 aangeven, fungeren als een krachtig ontstekingsremmend middel, maar ook zijn metabolieten.
Inflammatoire mediatoren en enzymen onderdrukt door Nrf2
Cytokines en Chemokines
Cytokinen zijn eiwitten en polypeptiden met een laag molecuulgewicht die door een verscheidenheid aan cellen worden uitgescheiden; ze reguleren celgroei, differentiatie en immuunfunctie, en zijn betrokken bij ontsteking en wondgenezing. Cytokinen omvatten interleukinen (IL's), interferonen, tumornecrosefactor (TNF), koloniestimulerende factor, chemokinen en groeifactoren. Sommige cytokines worden beschouwd als pro-inflammatoire mediatoren, terwijl andere ontstekingsremmende functies hebben. Blootstelling aan oxidatieve stress resulteert in de overproductie van cytokinen die oxidatieve stress veroorzaken in doelwitcellen. Verschillende pro-inflammatoire cytokines worden overgeproduceerd wanneer NF-? B wordt geactiveerd door oxidatieve stress. Bovendien veroorzaakt pro-inflammatoire oxidatieve stress verdere activering van NF-? B en de overproductie van cytokinen. Activering van het Nrf2 / ARE-systeem speelt een belangrijke rol bij het doorbreken van deze cyclus. Chemokinen zijn een familie van kleine cytokinen, waarvan de belangrijkste rol het begeleiden van de migratie van ontstekingscellen is. Ze functioneren voornamelijk als chemoattractanten voor leukocyten, monocyten, neutrofielen en andere effectorcellen.
Er is gemeld dat activering van Nrf2 LPS-geïnduceerde transcriptionele opregulatie van pro-inflammatoire cytokines, waaronder IL-6 en IL-1? [50]. IL-1? en de productie van IL-6 wordt ook verhoogd in Nrf2? /? muizen met door dextraansulfaat geïnduceerde colitis [51], [52]. Nrf2 remt de productie van stroomafwaarts IL-17 en andere ontstekingsfactoren Th1 en Th17, en onderdrukt het ziekteproces in een experimenteel model van multiple sclerose, auto-immuun encefalitis [53]. De Nrf2-afhankelijke anti-oxidantgenen HO-1, NQO-1, Gclc en Gclm blokkeren TNF-?, IL-6, monocyte chemo attractant proteïne-1 (MCP1), macrofaag inflammatoir proteïne-2 (MIP2) en inflammatoir bemiddelaars. Maar in het geval van Nrf2-knockout-muizen treedt het ontstekingsremmende effect niet op [54]. Peritoneale neutrofielen van Nrf2-knock-out muizen behandeld met LPS hebben significant hogere niveaus van cytokines (TNF-? En IL-6) en chemokines (MCP1 en MIP2) dan wild-type (WT) cellen [54]. In vitro onderdrukt de overdracht van het Nrf2-gen naar de gladde spiercellen van de aorta van mensen en konijnen de secretie van MCP1 [8], [55] en Nrf2-afhankelijke HO-1-expressie onderdrukt TNF -? - gestimuleerde NF-? B en MCP-1 secretie in endotheelcellen van de menselijke navelstrengader [56]. Deze bevindingen suggereren dat, als reactie op inflammatoire stimuli, opregulatie van Nrf2-signalering de overproductie van pro-inflammatoire cytokines en chemokines remt en ook de activering van NF-? B beperkt.
Cell Adhesion Molecules
Celadhesiemoleculen (CAM's) zijn eiwitten die binden met cellen of met de extracellulaire matrix. Ze bevinden zich op het celoppervlak en zijn betrokken bij celherkenning, celactivering, signaaltransductie, proliferatie en differentiatie. Onder de CAM's zijn ICAM-1 en VCAM-1 belangrijke leden van de immunoglobulinesuperfamilie. ICAM-1 is in lage concentraties aanwezig in leukocyt- en endotheelcelmembranen. Bij cytokinestimulatie neemt de concentratie aanzienlijk toe. ICAM-1 kan worden geïnduceerd door IL-1 en TNF en wordt tot expressie gebracht door het vasculaire endotheel, macrofagen en lymfocyten. Het is een ligand voor integrine, een receptor die op leukocyten wordt aangetroffen. Wanneer de ICAM-1-integrinebrug wordt geactiveerd, binden leukocyten zich aan endotheelcellen en migreren ze vervolgens naar subendotheliale weefsels [57]. VCAM-1 bemiddelt de adhesie van lymfocyten, monocyten, eosinofielen en basofielen aan vasculair endotheel en draagt bij aan de rekrutering van leukocyten, wat uiteindelijk leidt tot weefselschade als gevolg van oxidatieve stress. Nrf2 remt de promotoractiviteit van VCAM-1 [58]. Het Nrf2-gereguleerde stroomafwaartse gen HO-1 kan de expressie van E-selectine en VCAM-1 beïnvloeden, adhesiemoleculen geassocieerd met endotheelcellen [59]. De pulmonale expressie van verschillende CAM's zoals CD-14, TREM1, SELE, SELP en VCAM-1 zijn significant hoger in Nrf2? /? muizen dan in Nrf2 + / + muizen [60]. Nrf2 in menselijke aorta-endotheelcellen onderdrukken TNF -? - geïnduceerde VCAM-1-expressie en interfereren met TNF -? - geïnduceerde monocytische U937-celadhesie [8]. Overexpressie van Nrf2 remt ook TNF -? - geïnduceerde VCAM-1 genexpressie in menselijke microvasculaire endotheelcellen [61]. Het van nature voorkomende antioxidant 3-hydroxyanthranilzuur (HA), een van de metabolieten van l-tryptofaan die in vivo wordt gevormd langs de metabole route die bekend staat als de kynurenine-route tijdens ontsteking of infectie, blijkt HO-1-expressie te induceren en Nrf2 in de navelstreng van de mens te stimuleren. ader-endotheelcellen (HUVEC's). Nrf2-afhankelijke HO-1-expressie geïnduceerd door HA remt MCP-1-secretie, VCAM-1-expressie en NF-KB-activering geassocieerd met vasculaire schade en ontsteking bij atherosclerose [56]. Het antiproliferatieve en ontstekingsremmende synthetische chalcon-derivaat 2?, 4?, 6? -Tris (methoxymethoxy) chalcon remt ICAM-1, de pro-inflammatoire cytokine IL-1? En TNF-? expressie in colonweefsel van muizen behandeld met trinitrobenzeensulfonzuur [62]. Opwaartse regulatie van Nrf2 remt de TNF -? - geïnduceerde ICAM-1-expressie in humane retinale pigmentepitheelcellen die zijn behandeld met lycopeen [63]. Al deze onderzoeken suggereren dat Nrf2 een sleutelrol speelt in het ontstekingsproces door de migratie en infiltratie van ontstekingscellen naar ontstoken weefsel te reguleren.
Matrix metalloproteïnasen (MMP's)
MMP's zijn wijdverbreid aanwezig in de extracellulaire matrix en zijn betrokken bij fysiologische en pathologische processen zoals celproliferatie, migratie, differentiatie, wondgenezing, angiogenese, apoptose en tumormetastase. Er is gerapporteerd dat de Nrf2 / HO-1-as MMP-9 remt in macrofagen en MMP-7 in menselijke darmepitheelcellen, en dit is gunstig bij de behandeling van inflammatoire darmaandoeningen [62], [64]. Door UV-straling geïnduceerde huidbeschadiging is ernstiger bij Nrf2-knock-out dan bij WT-muizen en het MMP-9-niveau is significant hoger, wat aangeeft dat Nrf2 de MMP-9-expressie vermindert. Daarom wordt Nrf2 beschouwd als beschermend tegen UV-straling [65]. Een andere studie meldde ook dat de neerwaarts gereguleerde transcriptionele activering van MMP-9 bij tumorcelinvasie en -ontsteking wordt gereguleerd door remming van de NF-KB-signaleringsroute [66]. Bij traumatisch ruggenmergletsel neemt de NF-KB-signaleringsroute ook deel aan het reguleren van de mRNA-niveaus van MMP-9 [67]. Daarom wordt bij inflammatie de regulatie van MMP's direct beïnvloed door de Nrf2-route of indirect via de Nrf2-beïnvloede NF-? B-route.
Cyclooxygenase-2 (COX2) en induceerbare stikstofoxidesynthase (INOS)
Een reeks experimenten met Nrf2-knockout-muizen hebben hun cruciale rol bij ontstekingen en de regulatie van pro-inflammatoire genen zoals COX-2 en iNOS aangetoond. Voor het eerst hebben Khor et al. meldde verhoogde expressie van pro-inflammatoire cytokines zoals COX-2 en iNOS in de colonweefsels van Nrf2? /? muizen vergeleken met WT Nrf2 + / + muizen, wat aangeeft dat Nrf2 hun activiteit onderdrukt [51]. Een ander rapport over voorbehandeling met sulforafaan, een van de bekende Nrf2-activatoren die aanwezig zijn in kruisbloemige groenten, toonde het ontstekingsremmende effect aan van het remmen van de expressie van TNF- ?, IL-1 ?, COX-2 en iNOS op zowel het mRNA. en eiwitniveaus in primaire peritoneale macrofagen van Nrf2 + / + muizen vergeleken met die van Nrf2? /? muizen [68]. Evenzo vertoont de hippocampus van Nrf2-knockout-muizen met door LPS geïnduceerde ontsteking ook een hogere expressie van ontstekingsmarkers zoals iNOS, IL-6 en TNF-? dan WT-muizen [69]. Evenzo zijn Nrf2-knock-out-muizen overgevoelig voor de oxidatieve stress veroorzaakt door 1-methyl-4-fenyl-1,2,3,6-tetrahydropyridine en vertonen ze verhoogde mRNA- en eiwitniveaus van ontstekingsmarkers zoals COX-2, iNOS , IL-6 en TNF-? [70]. Bovendien, levers van Nrf2? /? muizen die werden uitgedaagd met een methionine- en choline-deficiënt dieet, hebben een ~ 5 keer hogere mRNA-expressie van Cox2 en iNOS dan die van WT-muizen op hetzelfde dieet, wat duidt op een ontstekingsremmende rol van Nrf2 [71]. Onlangs hebben Kim et al. toonde aan dat het fytochemische ethylpyruvaat zijn ontstekingsremmende en antioxidatieve effecten uitoefent door de expressie van iNOS via Nrf2-signalering in BV2-cellen te verminderen. Ze toonden aan dat ethylpyruvaat de nucleaire translocatie van Nrf2 induceert, wat uiteindelijk de interactie tussen p65 en p300 remt, wat leidt tot verminderde expressie van iNOS [72]. Bovendien activeert de carbazool-analoog LCY-2-CHO Nrf2 en veroorzaakt de nucleaire translocatie ervan, wat leidt tot de onderdrukking van COX2- en iNOS-expressie [73] in vasculaire gladde spiercellen van de aorta van de rat.
Paradoxale rol van Nrf2 in de regulering van NLRP3 iIflammasome-activiteit
De NLR-familie, pyrin-domein met 3 (NLRP3) inflammasoom is een multiproteïne-complex dat functioneert als een pathogeenherkenningsreceptor (PRR) en het brede scala aan microbiële, oxidatieve stresssignalen herkent, zoals pathogeen-geassocieerde moleculaire patronen (PAMP's), Damage- geassocieerde moleculaire patroonmoleculen (DAMPs) en ROS [74]. Het geactiveerde NLRP3-inflammasoom medieert de splitsing van caspase-1 en de uitscheiding van pro-inflammatoir cytokine interleukine-1? (IL-1?) Die uiteindelijk het proces van celdood induceert dat bekend staat als pyroptose en dat gastheren beschermt tegen een breed scala aan pathogenen [75]. Afwijkende activering van het inflammasoom wordt echter in verband gebracht met afwijkende eiwitvouwziekten zoals overdraagbare spongiforme encefalopathieën, de ziekte van Alzheimer, de ziekte van Parkinson en ook type 2 diabetes [76], kanker [77], jicht en atherosclerose [78].
Een recente observatie van de Rong Hu-groep over de associatie van Nrf2 met negatieve regulatie van inflammasoom onthulde dat Nrf2 de NQO1-expressie induceert die leidt tot de remming van NLRP3-inflammasoomactivering, caspase-1-splitsing en IL-1? generatie in macrofagen. Bovendien reguleerde een bekende Nrf2-activator, tert-butylhydrochinon (tBHQ), negatief de NLRP3-transcriptie door de ARE te activeren op een Nrf2-afhankelijke manier [79]. Naast de bovenstaande waarneming, is dezelfde groep ook onthuld dat dimethylfumaraat (DMF) DSS-geïnduceerde colitis voorkomt via de activerende Nrf2-signaalroute die betrokken is bij Nrf2-nucleaire translocatie en remming van NLRP3-inflammasoomassemblage [80].
Een reeks experimenten met natuurlijke en synthetische verbindingen hebben ook het remmende effect van Nrf2 op NLRP3-inflammasoomactivering onthuld. Zo heeft de behandeling van epigallocatechine-3-gallate (EGCG) bij lupus-nefritis-muizen aangetoond dat het de renale NLRP3-inflammasoomactivering vermindert die wordt gemedieerd door de Nrf2-signaalroute [81]. Evenzo remt citral (3,7-dimethyl-2,6-octadienal), een belangrijke actieve stof in een Chinees kruidengeneesmiddel Litsea cubeba, de NLRP3-inflammasoomactivering via de Nrf2-antioxidant-signaleringsroute in muismodel met versnelde en ernstige lupus nefritis (ASLN) [82]. Evenzo beschermde biochanine tegen LPS / GalN-geïnduceerde leverbeschadiging door de Nrf2-route te activeren en de NLRP3-inflammasoomactivering in mannelijke BALB / c-muizen te remmen [83]. Bovendien bleek mangiferine de expressie van Nrf2 en HO-1 op een dosisafhankelijke manier te reguleren en remde het LPS / D-GalN-geïnduceerde hepatische NLRP3, ASC, caspase-1, IL-1? en TNF-? uitdrukking [84].
Ondanks de negatieve regulatie van NLRP3 door Nrf2, activeert het ook de NLRP3- en AIM2-inflammasoomfunctie. Haitao Wen en collega's ontdekten dat Nrf2? /? muizenmacrofagen hebben de defecte activering van het NLRP3 en AIM2 Inflammasoom aangetoond, maar niet het NLRC4 inflammasoom [85]. Interessant is dat deze waarneming de onbekende functies van Nrf2 in de context van ontstekingsgerelateerde ziekten weergeeft; daarom is het erg belangrijk om verder te bestuderen om het mechanisme te onthullen waarin Nrf2 de inflammasoomfunctie activeert voordat het als een therapeutisch doelwit wordt beschouwd.
Onderdrukking van pro-inflammatoire cytokine transcriptie door Nrf2
Een zeer recent onderzoek op basis van chromatine-immunoprecipitatie (ChIP) -seq en ChIP-qPCR-resultaten in muizenmacrofagen onthulde dat Nrf2 bindt aan de promotorregio's van pro-inflammatoire cytokines zoals IL-6 en IL-1? en remt RNA Pol II-rekrutering. Als gevolg hiervan is RNA Pol II niet in staat de transcriptionele activering van IL-6 en IL-1 te verwerken? dat leidt uiteindelijk tot remming van genexpressie. Voor de eerste keer onthulde de groep van Masayuki Yamamoto het nieuwe mechanisme waarmee Nrf2 niet alleen zijn stroomafwaartse genen transactiveert via ARE's, maar ook de transcriptionele activering van specifieke genen met of zonder een ARE onderdrukt door de rekrutering van RNA Pol II te remmen [50].
Overspraak tussen Nrf2 en NF-? B Pathways
NF-? B is een eiwitcomplex dat verantwoordelijk is voor DNA-transcriptie dat in bijna alle soorten dierlijke cellen wordt aangetroffen en dat betrokken is bij verschillende processen zoals ontsteking, apoptose, de immuunrespons, celgroei en ontwikkeling. p65, een Rel-eiwit van de NF-? B-familie, heeft een transactiveringsdomein terwijl p50 geen heterodimerisatie met Rel-eiwit nodig heeft om transcriptie te activeren. Tijdens oxidatieve stress wordt I? B-kinase (IKK) geactiveerd en veroorzaakt de fosforylering van I? B, wat resulteert in de afgifte en nucleaire translocatie van NF-? B. NF-? B veroorzaakt de transcriptie van pro-inflammatoire mediatoren zoals IL-6, TNF- ?, iNOS, IL-1 en intracellulaire adhesie COX-2.
Abnormale regulatie van NF-? B is in verband gebracht met reumatoïde artritis, astma, inflammatoire darmaandoeningen en door Helicobacter pylori-infectie geïnduceerde gastritis [86]. Momenteel wordt aangenomen dat NF-kB-activiteit de Keapl / Nrf2 / ARE-signaleringsroute voornamelijk in drie aspecten beïnvloedt: ten eerste, Keap1 degradeert IKK? door ubiquitinatie, waardoor de activiteit van NF-? B wordt geremd [87]. Ten tweede induceert het ontstekingsproces ontstekingsmediatoren zoals COX2 afgeleid van het cyclopentenon prostaglandine 15d-PGJ2, een sterk elektrofiel dat reageert met Keap1 en Nrf2 activeert, waardoor gentranscriptie wordt geïnitieerd met gelijktijdige remming van NF-KB-activiteit [58], [88] ( Afb.3 A, B). Ten derde kan NF-? B worden gecombineerd met de competitieve Nrf2 transcriptionele co-activator CBP [89], [90] (Fig. 3 C, D).
Figuur 3 Overspraak tussen de Nrf2- en NF-? B-paden. (A) Keap1 stuurt de IKK naar CUL3-gemedieerde ubiquitinatie en proteasoomafbraak die uiteindelijk leidt tot de remming van NF-? B-fosforylering en dit mechanisme werkt ook als competitieve binding van Nrf2 en IKK met Keap1. (B) Oxidatieve stress activeert IKK die NF-? B fosforyleert, wat leidt tot de translocatie ervan naar de kern en activering van pro-inflammatoire cytokines zoals COX-2. Het eindproduct van COX-2, bekend als 15d-PGJ2, werkt als een inductor van Nrf2 dat uiteindelijk leidt tot de onderdrukking van oxidatieve stress. (C) Nrf2 bindt zich met zijn transcriptionele cofactor CBP samen met kleine Maf en andere transcriptionele machines om ARE-gestuurde genexpressie te initiëren. (D) Wanneer NF-? B op een competitieve manier aan CBP bindt, remt het de binding van CBP met Nrf2, wat leidt tot de remming van Nrf2-transactivering.
Aangenomen wordt dat de Nrf2- en NF-? B-signaleringsroutes een interactie aangaan om de transcriptie of functie van stroomafwaartse doeleiwitten te regelen. Ter rechtvaardiging van deze aanname laten veel voorbeelden zien dat directe of indirecte activering en remming optreedt tussen leden van de Nrf2- en NF-? B-routes (Fig. 4). Als reactie op LPS verhoogt Nrf2 knockdown significant de NF-? B transcriptionele activiteit en NF-? B-afhankelijke gentranscriptie, wat aantoont dat Nrf2 de NF-? B activiteit belemmert [60], [91]. Bovendien remt verhoogde expressie van Nrf2-afhankelijke stroomafwaartse HO-1 de NF-? B-activiteit. Wanneer prostaatkankercellen kortstondig worden blootgesteld aan β-tochoferylsuccinaat, een derivaat van vitamine E, wordt de HO-1-expressie opgereguleerd. De eindproducten van HO-1 remmen de nucleaire translocatie van NF-? B [92]. Deze in vivo studies suggereren dat Nrf2 de NF-KB-signaalpad negatief reguleert. LPS stimuleert NF-? B DNA-bindingsactiviteit en het niveau van de p65-subeenheid van NF-? B is significant hoger in nucleaire extracten uit de longen van Nrf2? /? dan van WT-muizen, wat een negatieve rol suggereert van Nrf2 in NF-? B-activering. Bovendien, Nrf2? /? muizenembryofibroblasten behandeld met LPS en TNF-? toon meer prominente NF-? B-activering veroorzaakt door IKK-activering en I? B-? afbraak [60]. En respiratoire syncytiële virusklaring is significant verminderd, terwijl NF-? B DNA-bindende activiteit is verhoogd in Nrf2? /? muizen vergeleken met WT-muizen [93]. Pristane-geïnduceerde lupus-nefritis in Nrf2? /? muizen die gelijktijdig worden behandeld met sulforafaan hebben ernstige nierbeschadiging en pathologische veranderingen, evenals verhoogde iNOS-expressie en NF-? B-activering in vergelijking met de WT, wat suggereert dat Nrf2 lupus-nefritis verbetert door de NF-? B-signaalroute te remmen en ROS op te ruimen [94 ]. NF-? B-activiteit treedt ook op wanneer cellen worden behandeld met een Nrf2-inductor samen met LPS en TNF- ?. Een synthetisch chalconderivaat remt bijvoorbeeld de door TNF -? - geïnduceerde NF-? B-activering zowel direct als indirect en gedeeltelijk door de inductie van HO-1-expressie in HT-29-cellen van het menselijke intestinale epitheel [62]. Onderdrukking van NF-? B-translocatie en DNA-bindende activiteit evenals de onderdrukking van iNOS-expressie in hepatocyten worden gevonden wanneer F344-ratten worden behandeld met 3H-1,2-dithiol-3-thion (D3T) [95]. Na gelijktijdige behandeling met sulforafaan en LPS, de LPS-geïnduceerde expressie van iNOS, COX-2 en TNF-? in Raw 264.7 worden macrofagen gedownreguleerd, wat suggereert dat sulforafaan een ontstekingsremmende werking heeft via remming van NF-B DNA-binding [96]. Hoewel er verschillende experimentele studies zijn gedaan om het verband tussen de Nrf2- en NF-? B-routes te verklaren, blijven de resultaten tegenstrijdig. Zowel positieve als negatieve voorschriften zijn gerapporteerd tussen Nrf2 en NF-KB [97]. Gewoonlijk activeren chemopreventieve elektrofielen 3H-1,2-dithiole-3-thione, sulforafaan en Triterpenoid CDDO-Me Nrf2 door NF-kB en de gedownoreguleerde genen [98], [99], [100] te remmen. Daarentegen is aangetoond dat verschillende agentia of aandoeningen zoals ROS, LPS, stroomafschuifspanning, geoxideerd LDL en sigarettenrook zowel Nrf2- als NF-KB-activiteit [97] verhogen. Bovendien hebben in vivo onderzoeken aangetoond dat de NF-KB-activiteit afneemt in levers geïsoleerd uit Nrf2? /? muizen en NF-? B-bindingsactiviteit is lager in Nrf2? /? dan in Nrf2 + / + muizen [101]. Echter, menselijke aorta-endotheelcellen die zijn behandeld met adenovirale vector Nrf2 remmen NF-? B stroomafwaartse genen zonder de activiteit van NF-? B te beïnvloeden [8].
Figuur 4 Regulerende lus van Nrf2 en NF-? B. De Nrf2-route remt de NF-? B-activering door de afbraak van I? B-? en het verhogen van HO-1-expressie en antioxiderende afweer die ROS en ontgiftende chemicaliën neutraliseren. Als resultaat wordt ROS-geassocieerde NF-? B-activering onderdrukt. Evenzo vermindert NF-? B-gemedieerde transcriptie Nrf2-activering door te verminderenZIJNTranscriptie van het gen en vrij CREB-bindend eiwit door te concurreren met Nrf2 voor CBP. Bovendien verhoogt NF-? B de rekrutering van histondeacetylase (HDAC3) naar het ARE-gebied en daarom wordt Nrf2-transcriptionele activering voorkomen.
De activering van de Nrf2-signaleringsroute speelt een belangrijke rol bij de expressie van enzymen en genen die betrokken zijn bij de ontgifting van reactieve oxidanten door de antioxidantcapaciteit van de cellen in het menselijk lichaam te verbeteren. Hoewel veel onderzoeksstudies vandaag beschikbaar zijn, zijn de reguleringsmechanismen bij de activering van Nrf2 niet volledig begrepen. Een mogelijke rol van de Nrf2-signaleringsroute bij de behandeling van ontstekingen is ook gevonden. Dr. Alex Jimenez DC, CCST Insight
De rol van Nrf2 bij inflammatoire ziekten
In vivo-onderzoeken hebben aangetoond dat Nrf2 een belangrijke rol speelt bij ontstekingsziekten die verschillende systemen beïnvloeden; deze omvatten gastritis, colitis, artritis, longontsteking, leverschade, hart- en vaatziekten, neurodegeneratieve ziekte en hersenschade. In deze onderzoeken, Nrf2? /? dieren vertoonden ernstigere symptomen van ontsteking en weefselschade dan WT-dieren. Daarom wordt aangenomen dat de Nrf2-signaleringsroute een beschermend effect heeft bij ontstekingsziekten. Intra-tracheale installatie van varkens-alvleesklier-elastase induceert chronische obstructieve longziekte, in het bijzonder emfyseem. Nrf2-deficiënte muizen zijn zeer gevoelig voor emfyseem en afgenomen expressie van HO-1, PrxI en het antiprotease-gen SLPI komen voor in alveolaire macrofagen. Nrf2 wordt beschouwd als een belangrijke regulator in het door macrofagen gemedieerde afweersysteem tegen longbeschadiging [102]. Nrf2-deficiënte muizen met emfyseem geïnduceerd door tabaksrookblootstelling gedurende 6 maanden vertonen toegenomen bronchoalveolaire ontsteking, opgereguleerde expressie van oxidatieve stressmarkers in longblaasjes en toegenomen alveolaire septale celapoptose, hetgeen suggereert dat Nrf2 werkt tegen tabak-geïnduceerd emfyseem door de verhoogde expressie van antioxidant genen [102], [103]. Bij een verstoring van Nrf2 vertonen allergeen-gemedieerde luchtwegontsteking en astma met ovalbumine-complex verhoogde luchtwegontsteking, luchtwegovergevoeligheid, hyperplasie van slijmbekercellen en hoge Th2-waarden in bronchoalveolaire lavage en splenocyten, terwijl de Nrf2-gemedieerde signaalroute eosinofilie van de luchtwegen beperkt , hypersecretie van slijmvliezen, en hyperreactiviteit van de luchtwegen, evenals het induceren van veel antioxiderende genen die de ontwikkeling van astma voorkomen [104]. Carrageenan injectie in de pleuraholte induceert pleuritis en 15d-PGJ2 accumulatie in Nrf2 ontstekingscellen is beperkt tot muis peritoneale macrofagen. Tijdens de vroege fase van ontsteking activeert 15d-PGJ2 Nrf2 en reguleert het ontstekingsproces via de inductie van HO-1 en PrxI. Een studie suggereerde ook dat COX-2 een ontstekingsremmend effect heeft in de vroege fase door de productie van 15d-PGJ2 [105]. Orale toediening van 1% dextransulfaatnatrium voor 1-week induceert colitis geassocieerd met histologische veranderingen die het verkorten van crypten en infiltratie van ontstekingscellen in colonweefsel omvatten. Om intestinale integriteit bij colitis te beschermen, zou Nrf2 een belangrijke rol kunnen spelen door pro-inflammatoire cytokines te reguleren en fase II-ontgiftende enzymen [51] te induceren. In een Nrf2-knock-out muismodel van LPS-geïnduceerde pulmonale sepsis, reguleert NF-? B-activiteit de invloed van inflammatoire cytokines zoals COX-2, IL-113, IL-6 en TNF? die essentieel zijn voor het initiëren en bevorderen van ontstekingen [60]. Nrf2 vermindert inflammatoire schade door deze ontstekingsfactoren te reguleren. In deze modellen van acute ontsteking vermindert de verhoogde regulatie van antioxidant-enzymen, pro-inflammatoire cytokines en mediatoren door de Nrf2-signaleringsroute de ontstekingsschade bij WT-dieren. Interessant is dat dit ook is gemeld in Nrf2-knockout-muizen waarbij de symptomen duidelijk zijn verergerd in vergelijking met WT-muizen.
Onderzoek naar Nrf2-afhankelijke ontstekingsremmende geneesmiddelen
Samenvattend hebben we experimenten besproken die aantonen dat de Nrf2-signaalroute een regulerende rol speelt in veel gebieden van ontsteking, dus Nrf2-afhankelijke ontstekingsremmende middelen zijn belangrijk voor de behandeling van ontstekingsziekten.
Planten zijn buitengewoon rijke bronnen van verbindingen die de Nrf2-transcriptiefactor activeren, wat leidt tot opwaartse regulatie van cytoprotectieve genen. Onlangs zijn verschillende onderzoeken uitgevoerd om de effecten van verschillende ontstekingsremmende middelen, meestal van plantaardige oorsprong, te onderzoeken. Curcumine is bijvoorbeeld het actieve ingrediënt van kurkuma en wordt ook in kleine hoeveelheden in gember aangetroffen; isothiocyanaten, in het bijzonder fenylisothiocyanaten zijn van broccoli, selderij en andere groenten; en anthocyanins zijn van bessen en druiven [124]. Studies hebben aangetoond dat al deze middelen niet alleen goede antioxidanten zijn, maar ook krachtige anti-inflammatoire effecten hebben via Nrf2-inductie [125], [126]. Daarom heeft de ontwikkeling van nieuwe ontstekingsremmende Nrf2-activatoren uit plantenextract veel belangstelling getrokken voor medisch onderzoek.
In de afgelopen jaren zijn er veel dierproeven uitgevoerd om de werking van deze verbindingen te bevestigen. Artesunate wordt voornamelijk gebruikt voor ernstige malaria, cerebrale malaria en reumatische auto-immuunziekten; het is ook effectief bij septische longbeschadiging. Artesunate activeert Nrf2- en HO-1-expressie, en de laatste vermindert de instroom van pro-inflammatoire cytokines en leukocyten in weefsel om ontsteking te voorkomen [127]. Isovitexine, gewonnen uit de rompen van Oryza sativa-rijst, wordt verondersteld ontstekingsremmende en antioxiderende eigenschappen te hebben; het speelt een beschermende rol tegen LPS-geïnduceerde acute longbeschadiging door de Nrf2 / HO-1-route te activeren en MAPK en NF-? B te remmen [128]. Fimasartan, een onlangs populaire angiotensine II-receptorantagonist die inwerkt op het renine-angiotensinesysteem, verlaagt de bloeddruk; het gebruik van fimasartan om muizen met chirurgisch geïnduceerde unilaterale ureterobstructie te behandelen vermindert oxidatieve stress, ontsteking en fibrose via opregulatie van Nrf2 en de antioxidantroute en remming van RAS en MAPK's [129]. Sappanon wordt wijd verspreid in Zuidoost-Azië, waar het wordt gebruikt als een anti-influenza, anti-allergisch en neuroprotectief medicijn; het activeert Nrf2 en remt NF-? B en kan dus nuttig zijn bij de behandeling van Nrf2- en / of NF-? B-gerelateerde ziekten [130]. Bixin gewonnen uit de zaden van Bixin orellana wordt gebruikt voor infectie- en ontstekingsziekten in Mexico en Zuid-Amerika; het vermindert inflammatoire mediatoren, alveolaire capillaire lekkage en oxidatieve schade op een Nrf2-afhankelijke manier om door ventilatie geïnduceerde longbeschadiging te verlichten en de normale longmorfologie te herstellen [131]. Andere plantaardige stoffen, zoals epigallocatechinegallaat, sulforafaan, resveratrol, lycopeen en groene thee-extract, hebben therapeutische effecten op ontstekingsziekten via de Nrf2-signaalroute [132], [133], [134]. Onlangs is gemeld dat een ander fytochemisch middel, eriodictyol, dat aanwezig is in citrusvruchten, ontstekingsremmende en antioxiderende effecten heeft op door cisplatine geïnduceerd nierletsel en sepsis-geïnduceerd acuut longletsel door Nrf2 te reguleren, NF-? B te remmen en de expressie van cytokinen in macrofagen [135], [136]. Talrijke fytochemicaliën zijn echter veelbelovend voor de preventie en behandeling van verschillende ziekten bij de mens, en sommige zijn al in het stadium van klinische proeven gekomen (tabel 2).
Deze plantverbindingen activeren de Nrf2-signaleringsroute hoofdzakelijk in de vorm van elektrofiele materialen die de cysteïne-residuen van Keap1 modificeren, wat leidt tot vrije nucleaire Nrf2-binding met de ARE, resulterend in activering van transcriptie van het overeenkomstige gen.
Sulforafaan en de effecten ervan op kanker, mortaliteit, veroudering, hersenen en gedrag, hartaandoeningen en meer
Isothiocyanaten zijn enkele van de belangrijkste plantaardige stoffen die u in uw dieet kunt krijgen. In deze video maak ik de meest uitgebreide case voor hen die ooit is gemaakt. Korte aandachtsspanne? Ga naar uw favoriete onderwerp door op een van de onderstaande tijdspunten te klikken. Volledige tijdlijn hieronder.
Belangrijkste secties:
00: 01: 14 - Kanker en sterfte
00: 19: 04 - Veroudering
00: 26: 30 - Hersenen en gedrag
00: 38: 06 - Laatste overzicht
00: 40: 27 - Dosis
Volledige tijdlijn:
00: 00: 34 - Introductie van sulforafaan, een belangrijk aandachtspunt van de video.
00: 01: 14 - Kruisbloemige groenteconsumptie en verlagingen van mortaliteit door alle oorzaken.
00: 02: 12 - Risico op prostaatkanker.
00: 02: 23 - Blaaskankerrisico.
00: 02: 34 - Longkanker bij rokers risico.
00: 02: 48 - Risico op borstkanker.
00: 03: 13 - Hypothetisch: wat als je al kanker hebt? (Interventie)
00: 03: 35 - Aannemelijk mechanisme dat de associatieve gegevens over kanker en mortaliteit stuurt.
00: 04: 38 - Sulforafaan en kanker.
00: 05: 32 - Dierlijk bewijs dat een sterk effect van extract van broccolispruit toont op de ontwikkeling van blaastumoren bij ratten.
00: 06: 06 - Effect van directe suppletie van sulforafaan bij prostaatkankerpatiënten.
00: 07: 09 - Bioaccumulatie van isothiocyanaatmetabolieten in daadwerkelijk borstweefsel.
00: 08: 32 - Remming van stamcellen van borstkanker.
00: 08: 53 - Geschiedenisles: brassica's werden vastgesteld met gezondheidseigenschappen zelfs in het oude Rome.
00: 09: 16 - het vermogen van Sulforaphane om de kankerverwekkende uitscheiding te verbeteren (benzeen, acroleïne).
00: 09: 51 - NRF2 als een genetische switch via antioxidantresponselementen.
00: 10: 10 - Hoe NRF2-activering de carcinogene excretie via glutathion-S-conjugaten verbetert.
00: 10: 34 - Spruitjes verhogen glutathion-S-transferase en verminderen DNA-schade.
00: 11: 20 - Broccolispruitendrank verhoogt de benzeenuitscheiding met 61%.
00: 13: 31 - Broccoli-spruithomogenaat verhoogt antioxiderende enzymen in de bovenste luchtwegen.
00: 15: 45 - Kruisbloemige groenteconsumptie en sterfte aan hart- en vaatziekten.
00: 16: 55 - Broccolispruitpoeder verbetert de bloedlipiden en het algemene risico op hartziekten bij type 2 diabetici.
00: 19: 04 - Begin van het verouderingsgedeelte.
00: 19: 21 - Met Sulforaphane verrijkte voeding verlengt de levensduur van kevers van 15 tot 30% (onder bepaalde omstandigheden).
00: 20: 34 - Belang van lage ontsteking voor een lang leven.
00: 22: 05 - Kruisbloemige groenten en broccoli-kiempoeder lijken een breed scala aan ontstekingsmarkers bij mensen te verminderen.
00: 24: 14 - Muisstudies suggereren dat sulforaphane de adaptieve immuunfunctie op oudere leeftijd zou kunnen verbeteren.
00: 25: 18 - Sulforafaan verbeterde de haargroei in een muismodel van kalend. Afbeelding bij 00: 26: 10.
00: 26: 30 - Begin van de sectie hersenen en gedrag.
00: 27: 18 - Effect van extract van broccolispruiten op autisme.
00: 27: 48 - Effect van glucoraphanine op schizofrenie.
00: 28: 17 - Begin van depressie discussie (plausibel mechanisme en studies).
00: 31: 21 - Muisstudie met behulp van 10 verschillende modellen van stress-geïnduceerde depressie laten sulforafaan zien, even effectief als fluoxetine (prozac).
00: 32: 00 - Onderzoek toont aan dat directe inname van glucoraphanine bij muizen even effectief is bij het voorkomen van depressies als gevolg van een sociaal nederlaagstressmodel.
00: 33: 01 - Begin van de sectie neurodegeneratie.
00: 33: 30 - Sulforafane en de ziekte van Alzheimer.
00: 33: 44 - Sulforafane en de ziekte van Parkinson.
00: 33: 51 - De ziekte van Sulforaphane en Hungtington.
00: 34: 43 - Begin van een traumatisch gedeelte over hersenletsel.
00: 35: 01 - Sulforafaan wordt geïnjecteerd onmiddellijk nadat TBI het geheugen verbetert (muisstudie).
00: 35: 55 - Sulforafaan en neuronale plasticiteit.
00: 36: 32 - Sulforafaan verbetert het leren in het model van type II diabetes bij muizen.
00: 37: 19 - Sulforafaan en duchenne spierdystrofie.
00: 37: 44 - Myostatinremming in spieratlanticellen (in vitro).
00: 38: 06 - Late video recapitulatie: sterfte en kanker, DNA-schade, oxidatieve stress en ontsteking, uitscheiding van benzeen, cardiovasculaire aandoeningen, type II diabetes, effecten op de hersenen (depressie, autisme, schizofrenie, neurodegeneratie), NRF2-route.
00: 40: 27 - Gedachten over het uitzoeken van een dosis broccoli of sulforafaan.
00: 41: 01 - Anekdotes over ontspruiten thuis.
00: 43: 14 - Op kooktemperaturen en sulforafaanactiviteit.
00: 43: 45 - Darmbacterieomzetting van sulforafaan uit glucoraphanine.
00: 44: 24 - Supplementen werken beter in combinatie met actieve myrosinase van groenten.
00: 44: 56 - Koken technieken en kruisbloemige groenten.
00: 46: 06 - Isothiocyanaten als goitrogens.
Conclusies
Momenteel heeft veel onderzoek zich gericht op de rol van de Nrf2 / Keap1 / ARE-signaleringsroute bij ontstekingen. Van de enzymen die door Nrf2 worden opgereguleerd, is HO-1 een van de representatieve stressrespons-enzymen. HO-1 heeft prominente ontstekingsremmende en antioxiderende eigenschappen. Over het algemeen reguleert de Nrf2-signaleringsroute ook negatief cytokines, chemokine-afgevende factoren, MMP's en andere inflammatoire mediatoren COX-2 en iNOS-productie, die direct of indirect de relevante NF-kB- en MAPK-routes en andere netwerken die ontstekingen beheersen beïnvloeden. Er wordt gesuggereerd dat de Nrf2- en NF-? B-signaleringsroutes een interactie aangaan om de transcriptie of functie van stroomafwaartse doeleiwitten te reguleren. Onderdrukking of inactivering van NF-? B-gemedieerde transcriptionele activiteit door Nrf2 vindt hoogstwaarschijnlijk plaats in de vroege fase van ontsteking, aangezien NF-? B de de novo synthese reguleert van een reeks pro-inflammatoire mediatoren. Er zijn echter nog enkele beperkingen in het onderzoek, zoals of er verbindingen zijn tussen Nrf2 en andere signaalroutes zoals JAK / STAT, de betekenis van de huidige Nrf2-activatoren afgeleid van natuurlijke plantaardige bronnen bij ontstekingen, en hoe de biologische activiteit kan worden verbeterd. en de doelgerichtheid van deze verbindingen verbeteren. Deze vereisen verdere experimentele validatie.
Bovendien kan de Nrf2-signaleringsroute> 600 genen [163] reguleren, waarvan> 200 coderen voor cytoprotectieve eiwitten [164] die ook worden geassocieerd met ontstekingen, kanker, neurodegeneratieve ziekten en andere belangrijke ziekten [165]. Groeiend bewijs dat suggereert dat de Nrf2-signaleringsroute in veel kankers is gedereguleerd, wat resulteert in afwijkende expressie van Nrf2-afhankelijke genenbatterij. Bovendien speelt ontsteking een belangrijke rol bij aan oxidatieve stress gerelateerde ziekten, vooral bij kanker. Toepassing van verschillende Nrf2-activators om de ontsteking tegen te gaan, kan resulteren in afwijkende expressie van Nrf2-stroomafwaartse genen die oncogenese en resistentie tegen chemo en/of radiotherapie induceren. Daarom kunnen zeer specifieke activatoren van Nrf2 worden ontwikkeld om de pleiotrope effecten ervan te minimaliseren. Verschillende activatoren van Nrf2 hebben een significante verbetering van de ontstekingsremmende functies bij oxidatieve stressgerelateerde ziekten laten zien. Het beste voorbeeld van Nrf2-activator dat is goedgekeurd door de FDA en veel wordt gebruikt voor de behandeling van ontstekingsziekten zoals multiple sclerose (MS), is dimethylfumaraat. Tecfidera� (geregistreerde naam van dimethylfumaraat door Biogen) effectief gebruikt voor de behandeling van terugkerende vormen van multiple sclerose bij een groot aantal patiënten [152]. De werkzaamheid van het gebruik van Nrf2-activators voor de behandeling van ontstekingsziekten vereist echter verdere validatie om de schadelijke effecten van Nrf2 te voorkomen. Daarom kan de ontwikkeling van therapieën voor de ontstekingsremmende activiteit die wordt gemedieerd door Nrf2 een aanzienlijke klinische impact hebben. Lopende studies van de Nrf2-signaleringsroute over de hele wereld zijn gewijd aan het ontwikkelen van zeer gerichte therapeutische middelen om de symptomen van ontsteking onder controle te houden en om kanker, evenals neurodegeneratieve en andere belangrijke ziekten te voorkomen en te behandelen.
Kortom, Nrf2 detecteert de niveaus van oxidatieve stress in het menselijk lichaam en helpt uiteindelijk de regulatie van antioxidanten en ontgiftende enzymen en genen te bevorderen. Omdat chronische ontsteking veroorzaakt door verhoogde niveaus van oxidatieve stress is geassocieerd met neurodegeneratieve aandoeningen, Nrf2 kan een essentiële rol spelen bij de behandeling van onder andere gezondheidsproblemen zoals de ziekte van Alzheimer. De reikwijdte van onze informatie is beperkt tot chiropractie en problemen met de wervelkolom. Om het onderwerp te bespreken, kunt u het aan Dr. Jimenez vragen of contact met ons opnemen via 915-850-0900 .
Extra topic discussie: Kniepijn verlichten zonder operatie
Kniepijn is een bekend symptoom dat kan optreden als gevolg van een verscheidenheid aan knieblessures en / of aandoeningen, waaronder sportverwondingen. De knie is een van de meest complexe gewrichten in het menselijk lichaam, omdat het bestaat uit de kruising van vier botten, vier ligamenten, verschillende pezen, twee menisci en kraakbeen. Volgens de American Academy of Family Physicians zijn de meest voorkomende oorzaken van kniepijn patellasubluxatie, patellatendinitis of jumper's knee en de ziekte van Osgood-Schlatter. Hoewel kniepijn het meest voorkomt bij mensen ouder dan 60 jaar, kan kniepijn ook voorkomen bij kinderen en adolescenten. Kniepijn kan thuis worden behandeld volgens de RICE-methoden, maar ernstige knieblessures kunnen onmiddellijke medische aandacht vereisen, inclusief chiropractische zorg. �
Neurodegeneratieve ziekten, zoals de ziekte van Alzheimer en de ziekte van Parkinson, treffen miljoenen mensen wereldwijd. Een verscheidenheid aan behandelingsopties is beschikbaar om de symptomen van verschillende neurodegeneratieve ziekten te behandelen, hoewel de resultaten vaak beperkt zijn. Onderzoeksstudies hebben aangetoond dat oxidatieve stress veroorzaakt door zowel interne als externe factoren een oorzaak kan zijn voor de ontwikkeling van neurodegeneratieve ziekten. De transcriptiefactor, Nrf2, is vastgesteld om te functioneren als een belangrijk verdedigingsmechanisme tegen oxidatieve stress. Het doel van het onderstaande artikel is om de effecten van te tonen Nrf2 over neurodegeneratieve aandoeningen.
Modulatie van Proteostasis door transcriptiefactor NRF2
Neurodegeneratieve ziekten zijn gekoppeld aan de accumulatie van specifieke eiwitaggregaten, wat duidt op een intieme verbinding tussen beschadigde hersenen en verlies van proteostase. Proteostase verwijst naar alle processen waarmee cellen de abundantie en vouwing van het proteoom regelen dankzij een breed netwerk dat de regulatie van signaleringsroutes, genexpressie en eiwitafbraaksystemen integreert. Deze review probeert de meest relevante bevindingen samen te vatten over de transcriptionele modulatie van proteostase die wordt uitgeoefend door de transcriptiefactor NRF2 (kernfactor (van erytroïde afgeleid 2) -achtig 2). NRF2 werd klassiek beschouwd als de belangrijkste regulator van de reactie van de antioxidantcellen, hoewel het momenteel opduikt als een sleutelcomponent van de transductiemachinerie om proteostase te behouden. Zoals we zullen bespreken, zou NRF2 kunnen worden beschouwd als een hub die noodsignalen verzamelt die zijn afgeleid van verkeerd gevouwen eiwitaccumulatie om een gecoördineerde en perdureerbare transcriptionele respons op te bouwen. Dit wordt bereikt door functies van NRF2 gerelateerd aan de controle van genen die betrokken zijn bij het onderhoud van de endoplasmatische reticulumfysiologie, het proteasoom en autofagie.
Nucleaire factor (van erytroïde afgeleid 2) -achtig 2 (NRF2) is een basisch-leucine-rits-eiwit dat tegenwoordig wordt beschouwd als een hoofdregulator van cellulaire homeostase. Het controleert de basale en stress-induceerbare expressie van meer dan 250-genen die een cis-werkende enhancer gemeen hebben die het antioxidantresponselement (ARE) [1], [2], [3], [4], [5] wordt genoemd. Deze genen nemen deel aan fase I-, II- en III-detoxificatiereacties, glutathion en peroxiredoxine / thioredoxinemetabolisme, NADPH-productie via de pentosefosfaatroute en malisch enzym, vetzuuroxidatie, ijzermetabolisme en proteostase [6]. Gezien deze brede cytoprotectieve functies, is het mogelijk dat een enkele farmacologische hit in NRF2 het effect van de belangrijkste boosdoeners van chronische ziekten, waaronder oxidatieve, inflammatoire en proteotoxische stress, zou kunnen verminderen. De rol van NRF2 in de modulatie van de antioxidantafweer en de resolutie van ontstekingen is in talloze studies behandeld (beoordeeld in [7]). Hier zullen we ons concentreren op zijn rol in proteostase, dat wil zeggen, de homeostatische controle van eiwitsynthese, vouwen, trafficking en degradatie. Er zullen voorbeelden worden gegeven in de context van neurodegeneratieve ziekten.
Verlies van Proteostase Invloeden NRF2 Activiteit bij neurodegeneratieve ziekten
Een algemeen kenmerk van neurodegeneratieve ziekten is het optreden van afwijkende aggregatie van sommige eiwitten. Zo worden verkeerd gevouwen eiwitaggregaten van β -synucleïne (β-SYN) aangetroffen in de ziekte van Parkinson (PD), β-amyloïde (A?) Plaques en hypergefosforyleerde TAU-neurofibrillaire klitten bij de ziekte van Alzheimer (AD), huntingtine (Htt) in Ziekte van Huntington (HD), superoxidedismutase 1 (SOD1) en TAR DNA-bindend eiwit 43 (TDP-43) bij amyotrofische laterale sclerose (ALS), prioneiwit (PrP) bij spongiforme encefalopathieën, enz. Eiwitaggregaten kunnen een impact hebben op verschillende cellulaire routes, die op hun beurt NRF2-niveaus en -activiteit kunnen beïnvloeden.
Verschillende lagen van regelgeving bepalen NRF2 activiteit goed
Onder fysiologische omstandigheden vertonen cellen lage NRF2-eiwitniveaus vanwege de snelle omzet. Als reactie op verschillende stimuli wordt NRF2-eiwit geaccumuleerd, komt de kern binnen en verhoogt de transcriptie van ARE-bevattende genen. Daarom is het beheer van NRF2-eiwitniveaus een belangrijk punt dat positieve en negatieve invoersignalen zou moeten integreren. Zoals we verder zullen bespreken, wordt NRF2 geactiveerd door diverse overlappende mechanismen om een snelle en efficiënte respons te orkestreren, maar aan de andere kant kan NRF2 worden geremd, waarschijnlijk in een tweede fase, om zijn reactie uit te schakelen.
Vanuit klassiek oogpunt wordt de activering van NRF2 beschouwd als een gevolg van de cellulaire respons op oxiderende of elektrofiele verbindingen. In dit opzicht speelt de ubiquitine E3-ligase-adapter Kelch-achtige ECH-geassocieerde proteïne 1 (KEAP1) een cruciale rol. Moleculaire details zullen verder worden behandeld in paragraaf 4.1. Kort gezegd fungeert KEAP1 als een redox-sensor vanwege kritische cysteïneresiduen die leiden tot NRF2-ubiquitinatie en proteasomale afbraak. Naast deze klassieke modulatie wordt NRF2 diepgaand gereguleerd door signaleringsgebeurtenissen. Het is inderdaad aangetoond dat verschillende kinasen NRF2 fosforyleren en reguleren. NRF2 kan bijvoorbeeld worden gefosforyleerd door mitogeen geactiveerde proteïnekinasen (MAPK's), hoewel de bijdrage ervan aan NRF2-activiteit onduidelijk blijft [8], [9], [10], [11]. PKA-kinase evenals enkele PKC-isozymen [12], CK2 [13] of Fyn [14] fosforyleren NRF2 die de stabiliteit ervan wijzigen. Eerder werk van onze groep meldde dat glycogeensynthase kinse-3? (GSK-3?) Remt NRF2 door nucleaire uitsluiting en proteasomale afbraak [15], [25], [26], [27], [28], [29], [30]. De moleculaire details worden besproken in paragraaf 4.1. Bovendien is NRF2 onderworpen aan andere soorten regelgeving. NRF2-acetylering door CBP / p300 verhoogt bijvoorbeeld de activiteit ervan [17], terwijl het wordt geremd door miR153, miR27a, miR142-5p en miR144 [16], of door methylering van cytosine-guanine (CG) -eilanden binnen de NRF2-promotor. [18].
Impact van eiwitaggregaten op NRF2-regelgevingsmechanismen
In deze sectie zullen we ons concentreren op de manier waarop accumulatie van verkeerd gevouwen proteïnen invloed kan hebben op NRF2-activiteit, waarbij sommige van de hierboven genoemde routes als illustratieve voorbeelden worden geboden. Ten eerste moeten we in overweging nemen dat de accumulatie van eiwitten nauw verbonden is met oxidatieve schade. Inderdaad, verkeerd gevouwen eiwitaccumulatie en aggregatie induceren abnormale productie van reactieve zuurstofspecies (ROS) uit mitochondria en andere bronnen [19]. Zoals hierboven vermeld, zal ROS redox-gevoelige cysteïnes van KEAP1 modificeren, wat leidt tot de afgifte, stabilisatie en nucleaire lokalisatie van NRF2.
Met betrekking tot proteïnopathieën, wordt een voorbeeld van ontregelde signaleringsgebeurtenissen die NRF2 kunnen beïnvloeden geleverd door de hyperactivering van GSK-3? in AD. GSK-3 ?, ook bekend als TAU-kinase, neemt deel aan de fosforylering van dit met microtubuli geassocieerde eiwit, wat resulteert in de aggregatie ervan, de vorming van neurofibrillaire klitten en onderbreking van axonaal transport (besproken in [20]). Aan de andere kant, GSK-3? vermindert dramatisch NRF2-niveaus en activiteit zoals hierboven vermeld. Hoewel niet algemeen aanvaard, stelt de amyloïde cascade voor dat giftige A? oligomeren verhogen GSK-3? activiteit samen met TAU-hyperfosforylering en neuronendood [21], [22]. Er zijn verschillende modellen om uit te leggen hoe A? is voorstander van GSK3-? activiteit. Bijvoorbeeld A? bindt zich aan de insulinereceptor en remt PI3K- en AKT-signaalroutes, die cruciaal zijn om GSK-3 te behouden? geïnactiveerd door fosforylering op zijn N-terminale Ser9-residu [23]. Aan de andere kant, extracellulaire A? interageert met Frizzled-receptoren, blokkeert WNT-signalering [24] en resulteert opnieuw in het vrijkomen van actief GSK-3 ?. Samengevat, A? accumulatie leidt tot abnormale hyperactivering van GSK-3?, waardoor een geschikte NRF2-respons wordt belemmerd.
Zoals in het volgende gedeelte wordt besproken, leiden verkeerd gevouwen eiwitten tot activering van PERK en MAPK's, die op hun beurt NRF2 [31], [8], [9], [10], [11] opvoeren. Bovendien is gedegreguleerde CBP / p300-activiteit gerapporteerd in verschillende proteïnopathieën [32] en een algemene afname van DNA-methylatie in AD-hersenen was ook [33] te zien, wat reden gaf om de relevantie van deze bevindingen in NRF2-regulering te onderzoeken.
Wij en anderen hebben bij necropsieën van PD- en AD-patiënten een toename van NRF2-eiwitniveaus en enkele van hun doelwitten waargenomen, zoals Heem-oxygenase 1 (HMOX1), NADPH-chinon-oxidase 1 (NQO1), p62, enz., Beide met immunoblot en door immunohistochemie [34], [35], [36], [37], [38], [39]. De opwaartse regulatie van NRF2 bij deze ziekten wordt geïnterpreteerd als een niet-succesvolle poging van de zieke hersenen om homeostatische waarden te herstellen. Een ander onderzoek gaf echter aan dat NRF2 voornamelijk gelokaliseerd is in het cytoplasma van AD-hippocampale neuronen, wat duidt op verminderde NRF2-transcriptieactiviteit in de hersenen [40]. Het is denkbaar dat de ongelijkheid van deze waarnemingen gerelateerd is aan veranderingen in de factoren die NRF2 beheersen langs de progressieve stadia van neurodegeneratie.
Drie belangrijke systemen dragen bij aan proteostase, namelijk de ongevouwen eiwitrespons (UPR), het ubiquitine-proteasoomsysteem (UPS) en autofagie. Vervolgens presenteren we bewijs om NRF2 te zien als een hub die noodsignalen verbindt die worden geactiveerd door eiwitaggregaten met de eiwitderivaten.
NRF2 neemt deel aan de Unfolded Protein Response (UPR)
NRF2-activering als reactie op de UPR
Oxidatieve eiwitvouwing in het ER wordt aangestuurd door een aantal verschillende routes, waarvan de meest geconserveerde het eiwit disulfide-isomerase (PDI) en het sulfhydryloxidase endoplasmatisch oxidoreductine 1 (ERO1? En ERO1? Bij zoogdieren) als disulfidedonor omvat. In het kort katalyseert PDI de vorming en verbreking van disulfidebindingen tussen cysteïneresiduen in eiwitten, terwijl ze vouwen, vanwege de reductie en oxidatie van zijn eigen cysteïne-aminozuren. PDI wordt gerecycled door de werking van het huishoudenzym ERO1, dat disulfidebindingen opnieuw in PDI introduceert [41]. Moleculaire zuurstof is de terminale elektronenacceptor van ERO1, die stoichiometrische hoeveelheden waterstofperoxide genereert voor elke geproduceerde disulfidebinding [42]. Peroxidasen (PRX4) en glutathionperoxidasen (GPX7 en GPX8) zijn sleutelenzymen om waterstofperoxide in het ER te verminderen. Wanneer dit oxido-reductieve systeem niet goed werkt, treedt er een abnormale ophoping van verkeerd gevouwen eiwitten op in het ER en wordt een reeks signalen genaamd de ongevouwen eiwitrespons (UPR) naar het cytoplasma en de kern gestuurd om de ER-homeostase te herstellen [43]. Er zijn drie membraangeassocieerde eiwitten geïdentificeerd voor het detecteren van ER-stress in eukaryoten: activerende transcriptiefactor 6 (ATF6), pancreas ER eIF2? kinase (PERK, ook dubbelstrengs RNA-geactiveerde proteïnekinase-achtige ER-kinase) en inositol-vereist kinase1 (IRE1). Het luminale domein van elke sensor is gebonden aan een chaperonne van 78 kDa genaamd glucose-gereguleerd eiwit (GRP78 / BIP). BIP dissocieert bij ER-stress om ongevouwen eiwitten te binden, wat leidt tot de activering van de drie sensoren [44].
NRF2 en zijn homoloog NRF1, ook gerelateerd aan de antioxidantrespons, nemen deel aan de transductie van de UPR naar de kern. In het geval van NRF1 bevindt dit eiwit zich op het ER-membraan en ondergaat het nucleaire translocatie na deglycosylatie of splitsing. Vervolgens leidt UPR-activering tot de verwerking van NRF1 en nucleaire accumulatie van het resulterende fragment in het nucleaire compartiment. Het vermogen om ARE-bevattende genen van dit NRF1-fragment te transactiveren, wordt echter nog steeds besproken [45].
Glover-Cutter en collega's toonden activatie van de NRF2-ortholoog van C. elegans, SKN-1, met verschillende ER-stressoren. Verhoogde SKN-1-expressie was afhankelijk van verschillende UPR-mediators, waaronder IRE1- of PERK-wormorthologen [46]. In PERK-deficiënte cellen leidt een verminderde eiwitsynthese tot ophoping van endogene peroxiden en daaropvolgende apoptose [47]. De effector die door PERK wordt gebruikt om de ER te beschermen tegen deze peroxiden, kan NRF2 zijn, omdat is gerapporteerd dat PERK NRF2 fosforyleert in Ser40, waardoor de degradatie ervan door KEAP1 [31] wordt voorkomen. De inductie van ASK1 speelt waarschijnlijk ook een rol in deze route door de TRAF2-gemedieerde kinase-actie van IRE1 [48]. Hoewel de rol van MAPK's in de regulatie van NRF2 nog steeds controversieel is, werd onlangs gesuggereerd dat de IRE1-TRAF2-ASK1-JNK-route NRF2 [49] (Fig. 1) zou kunnen activeren. Interessant is dat in C. elegans en menselijke cellen nieuw bewijs suggereert dat cysteïnesulfenylatie van IRE1-kinase bij zijn activeringslus IRE1-gemedieerde UPR remt en een p38-antioxidantreactie initieert die wordt aangestuurd door NRF2. De gegevens suggereren dat IRE1 een oude functie heeft als een cytoplasmatische sentinel die p38 en NRF2 [50] activeert.
Figuur 1 Regeling van NRF2 door de UPR. Accumulatie van ongevouwen of verkeerd gevouwen eiwitten in het endoplasmatisch reticulum kan de ongevouwen eiwitrespons (UPR) initiëren. Eerst wordt de chaperone BIP vrijgegeven uit het intraluminale domein van de ER-sensoren IRE1 en PERK om ongevouwen / verkeerd gevouwen eiwitten te binden. Dit maakt dimerisatie en trans-auto-fosforylatie van hun cytosolische domeinen mogelijk. PERK-activering resulteert in directe NRF2-fosforylatie bij Ser40, leidend tot NRF2-translocatie naar de kern en activering van doelgenen. IRE1-activatie induceert de rekrutering van TRAF2 gevolgd door ASK1 en JNK-fosforylatie en activering. Omdat van JNK is gemeld dat het NRF2 fosforyleert en activeert, is het redelijk om te denken dat IRE1-activering zou leiden tot verhoogde NRF2-activiteit.
Er zijn veel onderzoeken naar de inductie van de UPR uitgevoerd met de remmer van eiwitglycosylering tunicamycine. NRF2 lijkt essentieel te zijn voor de preventie van door tunicamycine geïnduceerde apoptotische celdood [31] en de activering ervan onder deze omstandigheden wordt aangedreven door de autofagische afbraak van KEAP1 [51]. Dienovereenkomstig verhoogde shRNA-gemedieerde silencing van NRF2-expressie in? TC-6-cellen, een muriene insulinoma? -Cellijn, de door tunicamycine geïnduceerde cytotoxiciteit significant en leidde tot een toename van de expressie van de pro-apoptotische ER-stressmarker CHOP10. Aan de andere kant verminderde NRF2-activering door 1,2-dithiol-3-thione (D3T) de cytotoxiciteit van tunicamycine en verzwakte de expressie van CHOP10 en PERK [52]. Interessant is dat reukneuronen die werden onderworpen aan systemische toediening van tunicamycine, NRF2 verhoogden in parallel met andere UPR-leden zoals CHOP, BIP, XBP1 [53]. Deze resultaten zijn uitgebreid naar in vivo studies, aangezien laterale ventriculaire infusie van tunicamycine bij ratten de expressie van PERK en NRF2 in de hippocampus induceerde, vergezeld van significante cognitieve gebreken, verhoogde TAU-fosforylering en Ap42-afzettingen [54].
NRF2 Up-reguleert sleutelgenen voor het onderhoud van de ER-fysiologie
Het ER-lumen heeft een overvloedige toevoer van GSH uit het cytosol nodig om de disulfide-chemie te behouden. NRF2 moduleert cruciale enzymen van het GSH-metabolisme in de hersenen, zoals cystine / glutamaattransport, β-glutamaatcysteïne synthetase (α-GS), glutamaat-cysteïne ligase katalytische en modulerende subeenheden (GCLC en GCLM), glutathionreductase (GR) en glutathionperoxidase (GPX) (besproken in [55]). De relevantie van NRF2 bij het in stand houden van GSH in de ER wordt ondersteund door de bevinding dat farmacologische of genetische activering van NRF2 resulteert in verhoogde GSH-synthese via GCLC / GCLM, terwijl het remmen van de expressie van deze enzymen door NRF2-knockdown een opeenhoping van beschadigde eiwitten in de ER die leiden tot de UPR-activering [56].
In C. elegans verschillende componenten van de UPR-doelgenen gereguleerd door SKN-1, waaronder Ire1, Xbp1 en Atf6. Hoewel NRF2 de expressie van verschillende peroxidase- (PRX) en glutathionperoxidase- (GPX) -genen opwaarts reguleert bij zoogdieren (besproken in [57]), is alleen GPX8 een bonafide ER-gelokaliseerd enzym dat het KDEL-ophaalsignaal herbergt [58]. Verlies van GPX8 veroorzaakt UPR-activering, lekkage van ERO1? -Afgeleide waterstofperoxide naar het cytosol en celdood. Waterstofperoxide afgeleid van ERO1? activiteit kan niet diffunderen van het ER naar het cytosol dankzij de gecoördineerde werking van GPX8 en PRX4 [59]. In dit opzicht onthulde een analyse van de antioxidant verdediging pathway-genexpressie-array met behulp van RNA uit wildtype en NRF2-nul muizenweefsel dat de expressie van GPX8 werd neerwaarts gereguleerd in de afwezigheid van NRF2 [60]. In lijn hiermee laten transcriptoomanalyse van patiëntenmonsters die lijden aan myeloproliferatieve neoplasmata, polycytemie of myelofibrose, ziekten die ook geassocieerd worden met oxidatieve stress en laaggradige chronische ontsteking, lagere expressieniveaus van zowel NRF2 als GPX8 laten zien in vergelijking met controlepersonen [61]. Er zijn nog geen studies die GPX8 specifiek betrekken bij de bescherming van de menselijke hersenen, maar een transcriptoomanalyse bij muizen duidt op een compenserende GPX8-toename als reactie op het Parkinson-toxine MPTP [62].
Impact van NRF2 op de UPR-ontregeling bij neurodegeneratieve aandoeningen
Defecten van PDI-enzymen en chronische activering van de UPR zouden vervolgens neurodegeneratie kunnen initiëren of versnellen. Door ziekte aangetaste neuronen, diermodellen van neurodegeneratieve ziekte evenals post-mortem menselijke weefsels vertoonden opwaartse regulatie van verschillende UPR-markers bij de meeste van deze aandoeningen. De wijziging van de PDI / UPR-route bij neurodegeneratieve aandoeningen is goed beoordeeld in [63], maar de volgende hoogtepunten uit postmortemmonsters in de hersenen moeten worden overwogen. PDI-niveaus zijn verhoogd in wirwar-dragende neuronen en in Lewy-lichamen van AD- en PD-patiënten, respectievelijk [64], [65]. PDI en ERP57 worden omhoog gereguleerd in CSF door ALS-patiënten en in hersenen van CJD-proefpersonen [66], [67], [68]. BIP, PERK, IRE1 en ATF6 zijn verhoogd in samples van patiënten met AD, PD of ALS [69], [70], [71], [67]. BIP, CHOP en XBP1 zijn verhoogd in post-mortem hersenstalen van HD [72], [73]. Bovendien werd up-regulatie van ERP57, GRP94 en BIP gevonden in cortexweefsels van CJD-patiënten [74]. Al met al laat dit bewijs zien dat de opeenhoping van verkeerd gevouwen eiwitten in het hersenparenchym leidt tot een schadelijke en chronische activering van de UPR. Interessant is dat er een recente studie is die activering van NRF2 door PERK in vroege AD linkt. In deze studie analyseerden de auteurs of door oxidatieve stress gemedieerde veranderingen in NRF2 en de UPR vroege gebeurtenissen in de AD-pathogenese kunnen vormen door menselijke perifere bloedcellen en een AD-transgeen muismodel in verschillende ziektestadia te gebruiken. Verhoogde oxidatieve stress en verhoogde pSer40-NRF2 werden waargenomen in mononucleaire cellen uit menselijk perifeer bloed geïsoleerd van individuen met milde cognitieve stoornissen. Bovendien rapporteerden zij een verminderde ER-calciumhomeostase en opwaartse gereguleerde ER-stressmarkers in deze cellen van personen met milde cognitieve stoornissen en milde AD [75].
Wederzijdse regulering van NRF2 en het ubiquitine-proteasoomsysteem (UPS)
De UPS moduleert NRF2-eiwitniveaus
De UPS neemt deel aan de afbraak van beschadigde of verkeerd gevouwen eiwitten en controleert de niveaus van sleutelregulerende moleculen in het cytosol en de kern. De centrale kern van dit systeem is een groot multisubunit-enzym dat een proteolytisch actief complex met de naam 20S bevat. Het kern-proteasoom van 20S breekt ongevouwen eiwitten af, maar de binding aan verschillende regulerende eiwitcomplexen verandert de substraatspecificiteit en -activiteit. De toevoeging van één of twee 19S-reguleringssubeenheden aan de 20S-kern vormt bijvoorbeeld het 26S-proteasoom en verandert de specificiteit voor native-gevouwen eiwitten [76], [77]. Proteasomale afbraak heeft covalente binding van ubiquitine nodig. Conjugatie van ubiquitine verloopt via een cascade-mechanisme met drie stappen. Eerst activeert het ubiquitine-activerende enzym E1 ubiquitine in een ATP-vereiste reactie. Vervolgens draagt één E2-enzym (ubiquitine-dragereiwit of ubiquitine-conjugatie-enzym) het geactiveerde ubiquitine van E1 over naar het substraat dat specifiek is gebonden aan een lid van de ubiquitine-eiwitligasfamilie, genaamd E3. Hoewel het exacte lot van het ubiquitinated-eiwit afhangt van de aard van de ubiquitineketen, resulteert dit proces doorgaans in de afbraak door het 26S-proteasoom [78].
De E3-ligase KEAP1 is de bekendste remmer van NRF2. Het mechanisme van KEAP1-regulering legt op elegante wijze uit hoe de NRF2-niveaus zich aanpassen aan oxidatie-schommelingen. Onder basale omstandigheden wordt nieuw gesynthetiseerd NRF2 gegrepen door het homodimer KEAP1, dat één NRF2-molecuul bindt aan twee aminozuursequenties met lage (aspartaat, leucine, glycine; DLG) en hoge (glutamaat, threonine, glycine, glutamaat; ETGE) affiniteit. De interactie met KEAP1 helpt om NRF2 te presenteren aan het CULLIN3 / RBX1-eiwitcomplex, wat resulteert in zijn ubiquitinatie en daaropvolgende proteasomale afbraak. Redox-modificatie van KEAP1 bemoeilijkt echter de presentatie van NRF2 aan de UPS vertegenwoordigd door CULLIN3 / RBX1. Als gevolg hiervan ontsnapt nieuw gesynthetiseerd NRF2 aan KEAP1-afhankelijke afbraak, hoopt het zich op in de kern en activeert het ARE-bevattende genen [79], [80], [81], [82].
De E3-ligase-adapter? -TrCP is ook een homodimeer dat deelneemt aan de signaleringsgebeurtenissen die verband houden met de fosforylering van NRF2 door GSK-3 ?. Dit kinase fosforyleert specifieke serineresiduen van NRF2 (aspartaat, serine, glycine, isoleucine serine; DSGIS) om een afbraakdomein te creëren dat vervolgens wordt herkend door? -TrCP en getagd voor proteasoomafbraak door een CULLIN1 / RBX1-complex. De identificatie van de specifieke aminozuren die worden gefosforyleerd door GSK-3? in deze degron werd uitgevoerd door een combinatie van plaatsgerichte mutagenese van het Neh6-domein, 2D-gelelektroforese [15], [26] en massaspectroscopie [83]. Bijgevolg remming van GSK-3? door zeer selectieve geneesmiddelen of siRNA's tegen GSK-3-isovormen resulteerde in een toename van NRF2-eiwitniveaus. Vergelijkbare resultaten werden gevonden met siRNA's tegen? -TrCP isovormen 1 en 2. Stabilisatie van NRF2 na GSK-3? remming trad op in KEAP1-deficiënte muizenembryofibroblasten en in een ectopisch tot expressie gebrachte NRF2-deletiemutant die de kritische ETGE-residuen ontbeerde voor binding met hoge affiniteit aan KEAP1, wat verder een KEAP1-onafhankelijke regulatie aantoont.
In de context van neurodegeneratieve ziekten kunnen we ons de modulatie van NRF2 door de UPS op twee verschillende manieren voorstellen. Enerzijds zou het KEAP1-systeem redoxonbalans detecteren die is afgeleid van verkeerd gevouwen eiwitaccumulatie, terwijl de GSK-3 /? - TrCP-as zou werken als een actieve deelnemer in signaleringstransductie die is veranderd door verlies van proteostase (Fig. 2).
Afbeelding 2 De UPS regelt de NRF2-niveaus goed. Onder homeostatische omstandigheden worden lage NRF2-niveaus gehandhaafd door de werking van de E3-ligase-adapters KEAP1 en? -TrCP. Links bindt NRF2 aan de Kelch-domeinen van een KEAP1-homodimeer via motieven met een lage (DLG) en een hoge (ETGE) affiniteit. Via zijn BTB-domein bindt KEAP1 tegelijkertijd aan een CULLIN3 / RBX1-complex, waardoor NRF2-ubiquitinatie en -degradatie door het 26S-proteasoom mogelijk wordt gemaakt. Bovendien GSK-3? fosforyleert Ser335- en Ser338-residuen van NRF2 om een afbraakdomein (DpSGIpSL) te creëren dat vervolgens wordt herkend door de ubiquitine-ligase-adapter? -TrCP en getagd voor proteasoomafbraak door een CULLIN3 / RBX1-complex. Juist, bij blootstelling aan reactieve zuurstofsoorten of elektrofielen worden kritische Cys-residuen in KEAP1 gemodificeerd, waardoor KEAP1 niet in staat is om efficiënt te interageren met NRF2 of CULLIN3 / RBX1 en dan verhoogt deze transcriptiefactor zijn halfwaardetijd en transcriptionele activiteit naar ARE-genen. Signaleringsroutes die resulteren in remming van GSK-3?, Zoals AKT-fosforylering op Ser9, resulteren in NRF2-verstoorde afbraak door het proteasoom, accumulatie en inductie van doelwitgenen.
NRF2 Verhoogt de UPS-activiteit via de transcriptionele controle van proteasoom-subeenheden
NRF2 reguleert de expressie van verschillende proteasoomsubeenheden opwaarts, waardoor de cel wordt beschermd tegen de ophoping van toxische eiwitten. Twintig proteasoom- en ubiquitinatiegerelateerde genen lijken te worden gereguleerd door NRF2, volgens een brede microarray-analyse van lever-RNA dat was opgezet met de NRF2-inductor D3T [84]. In een posterieure studie toonden dezelfde auteurs aan dat de expressie van de meeste subeenheden van het 26S-proteasoom tot drievoudig was verbeterd in levers van muizen die met D3T waren behandeld. De eiwitniveaus van de subeenheid en de proteasoomactiviteit werden gecoördineerd verhoogd. Er werd echter geen inductie gezien bij muizen waarbij de transcriptiefactor NRF2 was verstoord. De promotoractiviteit van de PSMB5 (20S) proteasoomsubeenheid nam toe met ofwel NRF2-overexpressie of behandeling met activatoren in embryonale fibroblasten van muizen, en ARE's werden geïdentificeerd in de proximale promotor van PSMB5 [85]. Farmacologische activering van NRF2 resulteerde in verhoogde expressieniveaus van representatieve proteasoomsubeenheden (PSMA3, PSMA6, PSMB1 en PSMB5) alleen in niet-senescente menselijke fibroblasten die functionele NRF2 bevatten [86]. NRF2-activering tijdens aanpassing aan oxidatieve stress resulteert in hoge expressie van de PSMB1 (20S) en PA28? subeenheden (of S11, proteasoomregulator) [87]. Bovendien toonden de resultaten van menselijke embryonale stamcellen aan dat NRF2 de expressie regelt van het proteasoomrijpingseiwit (POMP), een proteasoomchaperon, dat op zijn beurt de proliferatie moduleert van zichzelf vernieuwende menselijke embryonale stamcellen, drieledige differentiatie van de kiemlaag en cellulaire herprogrammering [ 88]. Alles bij elkaar geven deze onderzoeken aan dat NRF2 de expressie van sleutelcomponenten van de UPS opwaarts reguleert en daarom actief bijdraagt aan de klaring van eiwitten die anders giftig zouden zijn.
De NRF2-UPS-as in neurodegeneratieve ziekten
De rol van de UPS bij neurodegeneratieve ziekten is een gebied van intensief debat. Eerste studies rapporteerden verminderde proteasoomactiviteit in menselijke necropsieën van patiënten die waren getroffen door verschillende neurodegeneratieve ziekten. In andere onderzoeken met in vitro en in vivo benaderingen werd echter een onveranderde of zelfs verhoogde proteasoomactiviteit gevonden (beoordeeld in [89]). Een mogelijke verklaring voor dit verschil is dat de niveaus van de UPS-componenten kunnen veranderen tijdens ziekteprogressie en in verschillende hersenregio's, zoals is gesuggereerd voor NRF2-doelen.
Ondanks deze controverse moet worden opgemerkt dat up-regulatie van ARE-bevattende proteasoomgenen de UPS zal versterken door de klaring van toxische eiwitten in de hersenen te vergroten. Inderdaad, ablatie van NRF1, ook modulator van de antioxidantrespons, in neuronale cellen leidt tot verminderde proteasoomactiviteit en neurodegeneratie. Chromatin immunoprecipitatie-experimenten en transcriptionele analyse hebben aangetoond dat PSMB6 wordt gereguleerd door NRF1. Bovendien leidde genexpressieprofilering tot de identificatie van NRF1 als een belangrijke transcriptionele regulator van proteasoomgenen in neuronen, wat suggereert dat verstoringen in NRF1 kunnen bijdragen tot de pathogenese van neurodegeneratieve ziekten [90]. Interessant is dat NRF1 en zijn lange isovorm, TCF11 genoemd, ARE-bevattende proteasoomgenen naar boven konden reguleren bij proteasoominhibitie in een feedbacklus om te compenseren voor verminderde proteolytische activiteit [91], [92].
Wat NRF2 betreft, is er een verband tussen vermindering van de NRF2-, RPT6 (19 S) en PSMB5 (20 S) -niveaus in de middenhersenen van DJ-1-deficiënte muizen die zijn behandeld met de neurotoxine paraquat [93]. Bovendien geeft de natuurlijk voorkomende verbinding sulforafaan (SFN) een robuuster beeld van NRF2 als een cruciale modulator van de UPS. In vitro experimenten met murine neuroblastoom Neuro2A-cellen vertoonden een verhoogde expressie van de katalytische subeenheden van het proteasoom, evenals de peptidase-activiteiten in reactie op SFN. Deze drug beschermde cellen tegen waterstofperoxide gemedieerde cytotoxiciteit en eiwitoxidatie op een manier die afhankelijk is van de proteasoomfunctie [94]. Bovendien hebben Liu en collega's een reportermuis gebruikt om de UPS-activiteit te volgen in reactie op SFN in de hersenen. Deze muizen brengen alomtegenwoordig het groene fluorescentie-eiwit (GFP) tot expressie, gefuseerd aan een constitutief afbraaksignaal dat de snelle afbraak door de UPS (GFPu) bevordert. In de cerebrale cortex reduceerde SFN het niveau van GFPu met een parallelle toename van chymotrypsine-achtige (PSMB5), caspase-achtige (PSMB2) en trypsine-achtige (PSMB1) activiteiten van het 20 S-proteasoom. Bovendien onthulde de behandeling van Huntington-afgeleide cellen met SFN dat NRF2-activatie mHtt-afbraak en verminderde mHtt-cytotoxiciteit [95] verhoogde. Het belangrijkste mechanisme van SFN-actie is door inductie van NRF2 [96]. De specifieke bijdrage van NRF2 moet worden aangepakt met behulp van NRF2-nulsystemen in verdere studies.
Functionele verbinding tussen NRF2 en Macroautophagy
NRF2-eiwitniveaus worden gemoduleerd door de adapter-eiwit P62
Autofagie verwijst naar de afbraak van cytosolische componenten in lysosomen. Dit proces wordt gebruikt voor de opruiming van langlevende en verkeerd gevouwen eiwitten, evenals beschadigde organellen. Een eerste koppeling tussen NRF2 en autofagie werd voor het eerst waargenomen in verband met het adaptoreiwit p62, ook SQSTM1 [97], [98], [99], [100], [101] genoemd. Dit eiwit transporteert ubiquitinated proteïnen naar de proteasomale en lysosomale degradatiemachines en sequesteert beschadigde proteïnen tot aggregaten voorafgaand aan hun degradatie. P62 presenteert een ubiquitine-geassocieerd (UBA) domein, voor binding aan ubiquitinated eiwitten, en een LC3-interacterend gebied (LIR) voor integratie met het autofagosomale membraan via de autofagie receptor LC3.
Hoewel de p62-gemedieerde inductie van NRF2 en zijn doelwitgenen voor het eerst werd gerapporteerd in 2007 [102], werd het moleculaire mechanisme niet volledig begrepen tot de ontdekking van zijn interactie met KEAP1 [103], [98], [99], [100 ], [101]. Komatsu en collega's identificeerden een KEAP1-interacterende regio (KIR) in p62 die KEAP1 bond in dezelfde basisoppervlakzak als NRF2 en met een bindingsaffiniteit vergelijkbaar met het ETGE-motief in NRF2, wat duidt op concurrentie tussen p62 en NRF2. De fosforylering van Ser351 in het KIR-motief in p62 (349-DPSTGE-354) bleek zijn affiniteit voor KEAP1 te verhogen, concurreert met NRF2-binding en de accumulatie en transcriptionele activering van zijn doelwitgenen mogelijk te maken [98], [99]. Overexpressie van p62 leidde in feite tot verminderde NRF2-ubiquitinatie en daaruit voortvloeiende stabilisatie, evenals inductie van zijn doelwitgenen [104]. Van sommige kinasen is gesuggereerd dat ze deelnemen aan p62-fosforylering. Het zoogdierdoelwit van rapamycinecomplex 1 (mTORC1) kan worden geïmpliceerd, aangezien behandeling met de mTOR-remmer rapamycine de fosforylering van p62 en de neerwaartse regulatie van KEAP1 na arsenietbehandeling onderdrukte. Onlangs werd aangetoond dat TGF -? - geactiveerd kinase 1 (TAK1) ook p62 kan fosforyleren, wat de afbraak van KEAP1 en NRF2-up-regulatie verbetert. De auteurs van deze studie suggereren dat dit een manier is om cellulaire redoxtasis onder steady-state-omstandigheden te reguleren, aangezien TAK1-deficiëntie ROS opwaarts reguleert in afwezigheid van een exogeen oxidatiemiddel in verschillende muisweefsels, parallel met een verlaging van NRF2-eiwitniveaus [105 ].
Een p62-construct zonder het UBA-domein was nog steeds in staat om KEAP1 te binden, wat impliceert dat de interactie niet afhankelijk was van ubiquitinated KEAP1 [101]. De p62-homoloog in Drosophila melanogaster, Ref (2) genaamd, bevat echter geen KIR-motief en heeft geen directe interactie met DmKEAP1, hoewel het via het UBA-domein aan ubiquitinated DmKEAP1 kan binden. Bovendien kan DmKEAP1 rechtstreeks interageren met Atg8 (homoloog aan zoogdier LC3). KEAP1-deficiëntie resulteert in Atg8 en autofagie-inductie afhankelijk van de NRF2 ortholoog CncC en onafhankelijk van TFEB / MITF [106]. De relatie tussen NRF2 en autophagy lijkt echter te worden behouden, met de nadruk op de functionele relevantie ervan.
De inductie van NRF2 door p62 is het resultaat van zowel de concurrentie om KEAP1 te binden als de afbraak van KEAP1 in het lysosoom. Silencing van p62 met siRNA verdubbelde de KEAP1 halfwaardetijd parallel met een afname van NRF2 en zijn doelgenen [101]. In overeenstemming bleek ablatie van p62-expressie verhoogde niveaus van KEAP1 vergeleken met wildtype muizen. Zeer relevant, de toename in KEAP1-niveaus werd niet beïnvloed door proteasoomremmers, maar werd verlaagd onder verhongering-inducerende autofagie [107]. KEAP1 is zelfs aanwezig in zoogdiercellen in autofagische vesikels versierd met p62 en LC3 [99], [100], [103]. Al deze gegevens suggereren dat KEAP1 een substraat is van de macro-autofagische machines, maar dit probleem moet met meer detail worden geanalyseerd vanwege het bestaan van enkele controversiële resultaten. KEAP1-eiwitniveaus waren verhoogd bij Atg7-nul-muizen, een belangrijke effector van macroautofagie [107], maar farmacologische remming van macro-autofagie met torin1, E64 / pepstatine of bafilomycine kon KEAP1 [107], [100] niet accumuleren. Over het algemeen suggereren deze resultaten dat verhoogde p62 niveaus KEAP1 sequenderen in autofagische vacuolen en waarschijnlijk resulteren dit in KEAP1 autofagische degradatie die NRF2-activering mogelijk maakt (Fig. 3). Twee verschillende studies meldden dat de sulfinezuurreductasen SESTRINS in deze context een belangrijke rol spelen. SESTRIN 2 communiceert met p62, KEAP1 en RBX1 en faciliteert p62-afhankelijke afbraak van KEAP1 en NRF2-activering van doelgenen [108]. Een andere studie toonde aan dat SESTRIN 2 interageerde met ULK1 en p62, wat de fosforylering van p62 bij Ser403 bevorderde, wat degradatie van ladingeiwitten, waaronder KEAP1 [109], vergemakkelijkte.
Figuur 3 NRF2-niveaus worden gereguleerd door het adaptoreiwit p62. De fosforylering van Ser 351 in het KIR-motief van p62 (349-DPSTGE-354) door mTORC1, TAK1 of andere kinasen resulteert in verhoogde affiniteit voor binding aan KEAP1 als gevolg van de gelijkenis met het ETGE-motief in NRF2. Dientengevolge verplaatst gefosforyleerd p62 NRF2 en bindt KEAP1. Het LIR-motief in p62 maakt interactie met LC3 in het autofagosomale membraan mogelijk, zodat het p62-KEAP1-complex uiteindelijk wordt afgebroken in het lysosoom. Als gevolg hiervan kan NRF2 zich ophopen, naar de kern transloceren en de transcriptie van ARE-bevattende genen, inclusief p62, verhogen. Dit regulerende mechanisme biedt een duurzame NRF2-respons, omdat KEAP1 nieuw moet worden gesynthetiseerd om NRF2-activiteit te remmen.
Modulatie van macroautofagie-genen door NRF2
NRF2 reguleert de expressie van relevante genen voor macroautofagie, evenals voor de UPR en de UPS. Het eerste bewijsmateriaal kwam uit onderzoeken waarin werd aangetoond dat p62-expressie werd geïnduceerd na blootstelling aan elektrofielen, ROS en stikstofmonoxide [110], [111], [112]. Het inductiemechanisme werd enige jaren later beschreven met de bevinding dat p62 een functionele ARE in zijn genpromoter [99] bevat. In een recente studie werden verschillende andere functionele ARE's gevonden en gevalideerd volgens bioinformatica-analyse en ChIP-assays. Bovendien vertoonden muis embryonale fibroblasten en corticale neuronen van Nrf2-knockout muizen verlaagde p62-expressie, die kon worden gered met een NRF2 tot expressie brengend lentivirus. Evenzo verlaagde NRF2-deficiëntie de p62-niveaus in beschadigde neuronen van hippocampus muizen [36]. Daarom is gesuggereerd dat NRF2-activering de p62-niveaus verhoogt, resulterend in KEAP1-afbraak en bevordert verdere NRF2-stabilisatie in een positieve feedbacklus. Dit niet-canonieke mechanisme van NRF2-inductie vereist veranderingen in genexpressie en kan een relevante reactie zijn op langdurige cellulaire stress.
Het ladingherkenningseiwit NDP52 bleek transcriptioneel gereguleerd door NRF2. NDP52 werkt op een vergelijkbare manier als p62, herkent ubiquitinated eiwitten en interageert met LC3 via een LIR-domein, zodat ladingen in lysosomen worden afgebroken. Vijf vermeende ARE's werden gevonden in Ndp52-promotor-DNA-sequentie. Drie van hen werden geïdentificeerd met verschillende mutante constructen en ChIP-testen als onmisbaar voor NRF2-gemedieerde Ndp52-transcriptie [113]. Van belang was dat de mRNA-niveaus van Ndp52 afnamen in de hippocampus van Nrf2-knockout-muizen. Eén van deze sequenties werd ook gevalideerd in een onafhankelijke studie als een NRF2-gereguleerde ARE [36].
De rol van NRF2 in de modulatie van autofagie is echter niet beperkt tot de inductie van deze twee ladingherkenningseiwitten. Om meer inzicht te krijgen in de rol van NRF2 in de modulatie van extra autofagiegerelateerde genen, screende onze groep de chromatin immunoprecipitatiedatabase ENCODE voor twee eiwitten, MAFK en BACH1, die binden aan de NRF2-gereguleerde ARE's. Met behulp van een script dat is gegenereerd op basis van de consensus ARE-sequentie van JASPAR, hebben we verschillende vermeende ARE's geïdentificeerd in veel autofagie-genen. Twaalf van deze sequenties werden gevalideerd als met NRF2 gereguleerde ARE's in negen autofagie-genen, waarvan de expressie verminderde in muizenembryofibroblasten van Nrf2-knockout-muizen, maar die konden worden hersteld door een lentifirus dat NRF2 tot expressie brengt. Onze studie toonde aan dat NRF2 de expressie activeert van sommige genen die betrokken zijn bij verschillende stappen van het autofagische proces, inclusief autofagie-initiatie (ULK1), vrachtherkenning (p62 en NDP52), autofagosoomvorming (ATG4D, ATG7 en GABARAPL1), verlenging (ATG2B en ATG5 ) en autolysosoomklaring (ATG4D). Bijgevolg was autofagie flux in reactie op waterstofperoxide aangetast als NRF2 afwezig was [36].
Relevantie van NRF2-gemedieerde macroautofagie-genen-expressie in neurodegeneratieve aandoeningen
Gebleken is dat defecte autofagie een belangrijke rol speelt bij verschillende neurodegeneratieve aandoeningen [114] en ablatie van autofagie leidt tot neurodegeneratie bij muizen [115], [116]. Atg7-knock-out muizen onthulden dat autofagie-deficiëntie resulteert in accumulatie van p62 in ubiquitine-positieve inclusielichaampjes. KEAP1 werd gesekwestreerd in deze inclusielichamen, leidend tot NRF2-stabilisatie en inductie van doelgenen [103]. Belangrijk is dat overmatige accumulatie van p62 samen met ubiquitinated eiwitten is geïdentificeerd bij neurodegeneratieve aandoeningen, waaronder AD, PD en ALS [117]. In feite brachten neuronen die hoge niveaus van APP of TAU van AD-patiënten tot expressie brachten ook p62 en nucleaire NRF2 tot expressie, hetgeen hun poging suggereert om intraneuronale aggregaten af te breken via autofagie [36].
NRF2-deficiëntie verergert eiwitaggregatie in de context van AD. In feite worden verhoogde niveaus van gefosforyleerde en in sarkosyl onoplosbare TAU gevonden in Nrf2-knock-out muizen, hoewel er geen verschil in kinase- of fosfatase-activiteiten kon worden gedetecteerd in vergelijking met de wild-type achtergrond [113]. Belangrijk is dat werd aangetoond dat NDP52 co-lokaliseert met TAU in muriene neuronen en dat directe interactie tussen fosfo-TAU en NDP52 werd aangetoond door co-immunoprecipitatie-experimenten zowel in muizen als in AD-monsters, wat wijst op zijn rol bij de afbraak van TAU. Interessant is dat uitschakeling van NDP52, p62 of NRF2 in neuronen resulteerde in verhoogde fosfo-TAU [113], [118]. Bovendien werden verhoogde intraneuronale APP-aggregaten gevonden in de hippocampus van APP / PS1? E9-muizen wanneer NRF2 afwezig was. Dit correleerde met veranderde autofagie-markers, waaronder verhoogde fosfo-mTOR / mTOR- en fosfo-p70S6k / p70S6k-verhoudingen (indicatief voor autofagie-remming), verhoogde niveaus van pre-cathepsine D en een groter aantal multivesiculaire lichamen [119]. Bij muizen die menselijk APP (V717I) en TAU (P301L) tot expressie brachten, leidde NRF2-deficiëntie tot verhoogde niveaus van totaal en fosfo-TAU in de onoplosbare fractie en verhoogde intraneuronale APP-aggregaten, samen met verminderde neuronale niveaus van p62, NDP52, ULK1, ATG5 en GABARAPL1. Co-lokalisatie tussen het adaptereiwit p62 en APP of TAU was verminderd in afwezigheid van NRF2 [36]. Over het algemeen benadrukken deze resultaten het belang van NRF2 bij neuronale autofagie.
Verschillende transcriptiefactoren werken gecoördineerd om Proteostasis te moduleren
Onder steady-state omstandigheden wordt proteostase gecontroleerd via eiwit-eiwit-interacties en post-translationele modificaties die een snelle respons verkrijgen. Cellulaire aanpassing vereist echter de transcriptionele regulatie van de UPR-, UPS- en autofagie-genen. Gezien het feit dat zenuwcellen continu worden onderworpen aan laagwaardige toxische aanvallen, waaronder oxidatieve en proteotoxische stress, kan een versterking van proteostase geïnduceerd door transcriptionele modulatie helpen hersenafbraak te voorkomen.
In het geval van de UPR zal de activering van elk van de drie armen uiteindelijk resulteren in de transcriptionele inductie van bepaalde genen (beoordeeld in [43]). Een ATF6-afgeleid fragment (ATF6f) bindt bijvoorbeeld aan ER-stressresponselementen (ERSE) en induceert de expressie van verschillende genen, waaronder XBPI, BIP en CHOP. Bovendien leidt PERK-signalering tot de activering van de transcriptiefactor ATF4, die de expressie van meerdere UPR-gerelateerde genen en enkele anderen bestuurt, waaronder de NRF2-doelgenen Hmox1 en p62. Ten slotte resulteert IRE1-activering in het genereren van een actieve transcriptiefactor, gesplitste XBP1 (XBP1s), die de transcriptie van genen bestuurt die coderen voor eiwitten die betrokken zijn bij eiwitvouwing.
Aan de andere kant bleek NRF1 noodzakelijk voor proteasomale genexpressie in de hersenen, omdat Nrf1-knock-out muizen verminderde expressie van genen die coderen voor verschillende subeenheden van de 20S-kern, evenals het 19S-regulerende complex samen met verminderde proteasomale functie vertoonden [90 ]. Zowel NRF1 als NRF2 binden aan ARE-sequenties in de promotorregio's van zijn doelwitgenen, wat suggereert dat ze overlappende transcriptionele activiteiten hebben, hoewel ze verschillen in hun regulerende mechanismen en cellulaire lokalisatie [120].
Transcriptiefactoren van de Forkhead-box O (FOXO) -familie bepalen de expressie van meerdere autofagiegerelateerde genen. Vergelijkbaar met wat er gebeurt met NRF2, zijn er meerdere regelingslagen voor de activiteit van FOXO-leden, die kunnen worden geïnduceerd door voedings- of oxidatieve stress [121]. Ten slotte speelt de transcriptiefactor TFEB, die wordt beschouwd als de meesterregulator van lysosomale biogenese, een cruciale rol in de regulatie van autofagie onder voedingsstressomstandigheden. Aldus leidt remming van mTORC1 tot nucleaire translocatie van TFEB en inductie van de expressie van autofagie-genen [122].
Over het algemeen suggereert het bestaan van verschillende transcriptionele regulatoren van deze machines ook overspraak en gedeeltelijk overbodige mechanismen die proteostasis onder verschillende omstandigheden kunnen waarborgen. NRF2 kan daarom een relevante rol spelen in weefsels die hoge niveaus van oxidatieve stress ondersteunen. Bijvoorbeeld, oxidatieve stress-geïnduceerde NRF2 kan functioneren onder nutriëntrijke omstandigheden om transcriptie up-reguleren autophagy, vergelijkbaar met wat is gevonden voor TFEB onder uithongering omstandigheden. Bovendien functioneert het brein grotendeels onder nutriëntenrijke omstandigheden, waardoor NRF2 wordt voorgesteld als een relevant mechanisme om autofagie in neuronen te activeren.
Veelbelovend therapeutisch potentieel voor NRF2 bij proteïnopathieën
In de afgelopen paar jaar is er een grote vooruitgang geboekt in de kennis van de regelgevende rollen van de UPR, UPS en autofagie op NRF2-activiteit, evenals de reciproke NRF2-gemedieerde transcriptie van componenten van deze drie systemen. Daarom kunnen nieuwe therapeutische mogelijkheden ontstaan op basis van de exploitatie van NRF2 als een cruciale regulator van de klaring van eiwitten bij neurodegeneratieve aandoeningen.
Een belangrijke resterende vraag is echter of het nuttig of schadelijk zal zijn om NRF2-niveaus in de hersenen te verhogen. Analyse van epidemiologische gegevens kan een gedeeltelijk antwoord geven, omdat het aangeeft dat het NFE2L2-gen zeer polymorf is en dat sommige enkelvoudige nucleotide polymorfismen die in het regulerende gebied van de promotor worden aangetroffen, een reeks fysiologische variabiliteit in genexpressie op populatieniveau en sommige haplotypes kunnen opleveren. werden geassocieerd met een verminderd risico en / of vertraagd begin van AD, PD of ALS [123]. Bovendien, zoals besproken door Hayes en collega's [124], zou het NRF2-effect een U-vormige respons kunnen hebben, wat betekent dat te lage NRF2-niveaus kunnen resulteren in een verlies van cytoprotectie en verhoogde gevoeligheid voor stressoren, terwijl te veel NRF2 het homeostatische evenwicht naar een reductief scenario (reductieve stress), dat verkeerd vouwen en aggregeren van eiwitten zou bevorderen. Lage NRF2-spiegels in de hersenen ondersteunen het idee dat een lichte opwaartse regulatie voldoende kan zijn om onder pathologische omstandigheden een voordeel te behalen. In feite is de beschermende rol van farmacologische NRF2-gemedieerde activering van eiwitklaring aangetoond in verschillende neurodegeneratiecelculturen en in vivo-modellen.
SFN is een farmacologische NRF2-activator waarvan is aangetoond dat het proteasomale en autofagie genexpressie induceert [95], [36]. Interessant genoeg toonden Jo en collega's aan dat SFN de niveaus van gefosforyleerde TAU verlaagde en Beclin-1 en LC3-II verhoogde, wat suggereert dat NRF2-activering de afbraak van dit toxische eiwit door autofagie kan vergemakkelijken [113]. Bovendien werd de afbraak van mHtt versterkt met SFN, en dit werd teruggedraaid door het gebruik van MG132, wat wijst op proteasomale afbraak van dit toxische eiwit [95]. Autofagie-gemedieerde afbraak van fosfo- en onoplosbaar-TAU werd gerapporteerd met het organische flavonoïde fisetine. Deze verbinding was in staat autofagie te induceren door tegelijkertijd de activering en nucleaire translocatie van zowel TFEB als NRF2 te bevorderen, samen met enkele van zijn doelwitgenen. Deze reactie werd voorkomen door TFEB of NRF2 silencing [125]. Bott en collega's rapporteerden gunstige effecten van een gelijktijdige NRF2-, NRF1- en HSF1-activator op eiwittoxiciteit bij spinale en bulbaire spieratrofie, een neurodegeneratieve aandoening veroorzaakt door expansie van polyglutamine-coderende CAG-herhalingen waarin eiwitaggregaten aanwezig zijn [126]. Het potentieel van NRF2-activering voor de behandeling van neurodegeneratieve aandoeningen is aangetoond met de goedkeuring van BG-12, de orale formulering van de NRF2-inductor dimethylfumaraat (DMF), voor de behandeling van multiple sclerose [127], [128]. Het succes van DMF met auto-immuunziekten met een sterke inflammatoire component suggereert dat neurodegeneratieve ziekten baat kunnen hebben bij herpositionering van dit medicijn. In een recente preklinische studie van een β -synucleinopathie-model van PD, bleek DMF neuroprotectief te zijn, gedeeltelijk als gevolg van de inductie van autofagie [129]. Studies die gunstige effecten van NRF2 op neurodegeneratie rapporteren, maar niet focussen op het effect ervan op de eiwitklaring, zijn zelfs overvloediger (voor een uitgebreid overzicht, zie [7]). Dit is behoorlijk relevant, omdat het de meerdere schadelijke processen benadrukt die gelijktijdig kunnen worden aangevallen door een enkele treffer in NRF2, waaronder ook oxidatieve stress, neuro-inflammatie of mitochondriale disfunctie. Er zal echter toekomstig werk nodig zijn om definitief te bepalen of farmacologische activering van NRF2 een geldige strategie kan zijn om de afbraak van toxische eiwitten in de hersenen te vergemakkelijken.
Zoals eerder uitgelegd, verergerde GSK-3? activiteit werd gerapporteerd bij neurodegeneratieve ziekten en er is gespeculeerd dat de daaruit voortvloeiende NRF2-reductie gedeeltelijk verantwoordelijk kan zijn voor het schadelijke resultaat. Onder deze pathologische omstandigheden zouden GSK-3-remmers ook kunnen samenwerken om NRF2-niveaus en proteostase te verhogen. De gunstige effecten van GSK-3-remmers zijn gerapporteerd in verschillende modellen van neurodegeneratie en, interessanter, GSK-3-repressie bleek de niveaus van toxische eiwitten te verlagen [130], [131], [132], [133]. Hoewel er nog geen directe verbanden zijn waargenomen tussen GSK-3-remming en NRF2-transcriptionele regulatie van genen die proteostase bevorderen, is het redelijk om te speculeren dat neerwaartse regulatie van GSK-3-activiteit zou resulteren in verhoogde NRF2-niveaus, wat uiteindelijk zal resulteren in versterkte proteostase.
De transcriptionele activiteit van NRF2 en het cellulaire vermogen om proteostase te behouden, nemen af met de leeftijd, de belangrijkste risicofactor voor de ontwikkeling van neurodegeneratieve ziekten. Het is redelijk om te denken dat de versterking van NRF2 en bijgevolg proteostase op zijn minst de accumulatie van eiwitaggregaten en neurodegeneratie zou vertragen. Inderdaad, behandeling van menselijke senescente fibroblasten met 18p-glycyrrhetinezuur (18p-GA) triterpenoïde bevorderde NRF2-activering, wat leidde tot proteasoominductie en verlengde levensduur. Deze studie suggereert dat farmacologische activering van NRF2 zelfs op latere leeftijd mogelijk is [86]. Bovendien toonde een latere studie aan dat deze verbinding SKN-1 en proteasoomactivering bij C.elegans bemiddelde met gunstige effecten op de progressie van AD in relevante nematodemodellen [134].
Alles bij elkaar genomen lijkt NRF2-gemedieerde inductie van proteostase-gerelateerde genen gunstig te zijn in verschillende proteïnopathieën.
Sulforafaan en de effecten ervan op kanker, mortaliteit, veroudering, hersenen en gedrag, hartaandoeningen en meer
Isothiocyanaten zijn enkele van de belangrijkste plantaardige stoffen die u in uw dieet kunt krijgen. In deze video maak ik de meest uitgebreide case voor hen die ooit is gemaakt. Korte aandachtsspanne? Ga naar uw favoriete onderwerp door op een van de onderstaande tijdspunten te klikken. Volledige tijdlijn hieronder.
Belangrijkste secties:
00: 01: 14 - Kanker en sterfte
00: 19: 04 - Veroudering
00: 26: 30 - Hersenen en gedrag
00: 38: 06 - Laatste overzicht
00: 40: 27 - Dosis
Volledige tijdlijn:
00: 00: 34 - Introductie van sulforafaan, een belangrijk aandachtspunt van de video.
00: 01: 14 - Kruisbloemige groenteconsumptie en verlagingen van mortaliteit door alle oorzaken.
00: 02: 12 - Risico op prostaatkanker.
00: 02: 23 - Blaaskankerrisico.
00: 02: 34 - Longkanker bij rokers risico.
00: 02: 48 - Risico op borstkanker.
00: 03: 13 - Hypothetisch: wat als je al kanker hebt? (Interventie)
00: 03: 35 - Aannemelijk mechanisme dat de associatieve gegevens over kanker en mortaliteit stuurt.
00: 04: 38 - Sulforafaan en kanker.
00: 05: 32 - Dierlijk bewijs dat een sterk effect van extract van broccolispruit toont op de ontwikkeling van blaastumoren bij ratten.
00: 06: 06 - Effect van directe suppletie van sulforafaan bij prostaatkankerpatiënten.
00: 07: 09 - Bioaccumulatie van isothiocyanaatmetabolieten in daadwerkelijk borstweefsel.
00: 08: 32 - Remming van stamcellen van borstkanker.
00: 08: 53 - Geschiedenisles: brassica's werden vastgesteld met gezondheidseigenschappen zelfs in het oude Rome.
00: 09: 16 - het vermogen van Sulforaphane om de kankerverwekkende uitscheiding te verbeteren (benzeen, acroleïne).
00: 09: 51 - NRF2 als een genetische switch via antioxidantresponselementen.
00: 10: 10 - Hoe NRF2-activering de carcinogene excretie via glutathion-S-conjugaten verbetert.
00: 10: 34 - Spruitjes verhogen glutathion-S-transferase en verminderen DNA-schade.
00: 11: 20 - Broccolispruitendrank verhoogt de benzeenuitscheiding met 61%.
00: 13: 31 - Broccoli-spruithomogenaat verhoogt antioxiderende enzymen in de bovenste luchtwegen.
00: 15: 45 - Kruisbloemige groenteconsumptie en sterfte aan hart- en vaatziekten.
00: 16: 55 - Broccolispruitpoeder verbetert de bloedlipiden en het algemene risico op hartziekten bij type 2 diabetici.
00: 19: 04 - Begin van het verouderingsgedeelte.
00: 19: 21 - Met Sulforaphane verrijkte voeding verlengt de levensduur van kevers van 15 tot 30% (onder bepaalde omstandigheden).
00: 20: 34 - Belang van lage ontsteking voor een lang leven.
00: 22: 05 - Kruisbloemige groenten en broccoli-kiempoeder lijken een breed scala aan ontstekingsmarkers bij mensen te verminderen.
00: 24: 14 - Muisstudies suggereren dat sulforaphane de adaptieve immuunfunctie op oudere leeftijd zou kunnen verbeteren.
00: 25: 18 - Sulforafaan verbeterde de haargroei in een muismodel van kalend. Afbeelding bij 00: 26: 10.
00: 26: 30 - Begin van de sectie hersenen en gedrag.
00: 27: 18 - Effect van extract van broccolispruiten op autisme.
00: 27: 48 - Effect van glucoraphanine op schizofrenie.
00: 28: 17 - Begin van depressie discussie (plausibel mechanisme en studies).
00: 31: 21 - Muisstudie met behulp van 10 verschillende modellen van stress-geïnduceerde depressie laten sulforafaan zien, even effectief als fluoxetine (prozac).
00: 32: 00 - Onderzoek toont aan dat directe inname van glucoraphanine bij muizen even effectief is bij het voorkomen van depressies als gevolg van een sociaal nederlaagstressmodel.
00: 33: 01 - Begin van de sectie neurodegeneratie.
00: 33: 30 - Sulforafane en de ziekte van Alzheimer.
00: 33: 44 - Sulforafane en de ziekte van Parkinson.
00: 33: 51 - De ziekte van Sulforaphane en Hungtington.
00: 34: 43 - Begin van een traumatisch gedeelte over hersenletsel.
00: 35: 01 - Sulforafaan wordt geïnjecteerd onmiddellijk nadat TBI het geheugen verbetert (muisstudie).
00: 35: 55 - Sulforafaan en neuronale plasticiteit.
00: 36: 32 - Sulforafaan verbetert het leren in het model van type II diabetes bij muizen.
00: 37: 19 - Sulforafaan en duchenne spierdystrofie.
00: 37: 44 - Myostatinremming in spieratlanticellen (in vitro).
00: 38: 06 - Late video recapitulatie: sterfte en kanker, DNA-schade, oxidatieve stress en ontsteking, uitscheiding van benzeen, cardiovasculaire aandoeningen, type II diabetes, effecten op de hersenen (depressie, autisme, schizofrenie, neurodegeneratie), NRF2-route.
00: 40: 27 - Gedachten over het uitzoeken van een dosis broccoli of sulforafaan.
00: 41: 01 - Anekdotes over ontspruiten thuis.
00: 43: 14 - Op kooktemperaturen en sulforafaanactiviteit.
00: 43: 45 - Darmbacterieomzetting van sulforafaan uit glucoraphanine.
00: 44: 24 - Supplementen werken beter in combinatie met actieve myrosinase van groenten.
00: 44: 56 - Koken technieken en kruisbloemige groenten.
00: 46: 06 - Isothiocyanaten als goitrogens.
De nucleaire factor van erytroïde afkomstige 2 (NF-E2) -gerelateerde factor 2, ook wel bekend als Nrf2, is een transcriptiefactor die de expressie van een verscheidenheid aan antioxidanten en ontgiftende enzymen reguleert. Onderzoeksstudies hebben ook zijn rol in het beheersen van oxidatieve stress aangetoond. De meeste neurodegeneratieve ziekten, zoals de ziekte van Alzheimer en de ziekte van Parkinson, worden gekenmerkt door oxidatieve stress en chronische ontsteking, de gemeenschappelijke doelen van Nrf2 behandelingsbenaderingen. Dr. Alex Jimenez DC, CCST Insight
Slotopmerkingen
Transcriptiefactor NRF2 orkestreert een proteostatische respons door het waarnemen en moduleren van veranderingen in de UPR, UPS en autofagie (figuur 4). Bijgevolg is aangetoond dat het ontbreken van NRF2 de proteïne-pathologie verergert, wat suggereert dat NRF2 noodzakelijk is voor optimale eiwitklaring. Alles bij elkaar kunnen we speculeren dat NRF2 een interessant therapeutisch doelwit kan zijn voor proteïnopathieën.
Figuur 4 NRF2 als een hub die proteotoxische afgeleide noodsignalen verbindt met een beschermende transcriptionele respons. De opeenhoping van ongevouwen / verkeerd gevouwen eiwitten zal leiden tot de activering van de ongevouwen eiwitrespons (UPR) in het ER. Activering van PERK of MAPK kan resulteren in de transcriptionele inductie van de ER-residente Gpx8 en verschillende enzymen die de GSH-niveaus reguleren, cruciaal om correcte eiwitvouwing te verzekeren. Eiwitaggregaten remmen proteasoomactiviteit (UPS), waardoor NRF2-degradatie waarschijnlijk wordt vermeden. Van NRF2 is aangetoond dat het de transcriptie van Psma3-, Psma6-, Psmb1-, Psmb5- en Pomp-genen specifiek moduleert. Verschillende andere subeenheden werden opwaarts gereguleerd op een NRF2-afhankelijke manier als reactie op D3T, waardoor waarschijnlijk de lijst van proteasoom-subeenheden die door NRF2 worden gereguleerd wordt vergroot. Autofagie is de belangrijkste route voor de afbraak van eiwitaggregaten. Autofagie reguleert ook NRF2, waarbij deze afbraakroute wordt verbonden met NRF2 transcriptionele inductie van p62, Ndp52, Ulk1, Atg2b, Atg4c, Atg5, Atg7 en Gabarapl1.
Volgens het artikel hierboven, terwijl de symptomen van neurodegeneratieve aandoeningen kunnen worden behandeld via een verscheidenheid aan behandelingsopties, hebben onderzoeken aangetoond dat Nrf2-activering een nuttige behandelaanpak kan zijn. Omdat Nrf2-activatoren richten zich op brede ziektemechanismenkunnen alle neurodegeneratieve ziekten profiteren van het gebruik van de Nrf2-transcriptiefactor. De bevindingen van Nrf2 hebben een revolutie teweeggebracht in de behandeling van neurodegeneratieve ziekten. De reikwijdte van onze informatie is beperkt tot chiropractie en problemen met de wervelkolom. Om het onderwerp te bespreken, kunt u het aan Dr. Jimenez vragen of contact met ons opnemen via 915-850-0900 .
Extra topic discussie: Kniepijn verlichten zonder operatie
Kniepijn is een bekend symptoom dat kan optreden als gevolg van een verscheidenheid aan knieblessures en / of aandoeningen, waaronder sportverwondingen. De knie is een van de meest complexe gewrichten in het menselijk lichaam, omdat het bestaat uit de kruising van vier botten, vier ligamenten, verschillende pezen, twee menisci en kraakbeen. Volgens de American Academy of Family Physicians zijn de meest voorkomende oorzaken van kniepijn patellasubluxatie, patellatendinitis of jumper's knee en de ziekte van Osgood-Schlatter. Hoewel kniepijn het meest voorkomt bij mensen ouder dan 60 jaar, kan kniepijn ook voorkomen bij kinderen en adolescenten. Kniepijn kan thuis worden behandeld volgens de RICE-methoden, maar ernstige knieblessures kunnen onmiddellijke medische aandacht vereisen, inclusief chiropractische zorg. �
Oxidatieve stress wordt beschreven als celbeschadiging veroorzaakt door vrije radicalen, of onstabiele moleculen, die uiteindelijk de gezonde functie kunnen beïnvloeden. Het menselijk lichaam creëert vrije radicalen om bacteriën en virussen te neutraliseren, maar externe factoren, zoals zuurstof, vervuiling en straling, kunnen vaak ook vrije radicalen produceren. Oxidatieve stress is in verband gebracht met tal van gezondheidsproblemen.
Oxidatieve stress en andere stressoren zetten interne beschermende mechanismen in werking die kunnen helpen bij het reguleren van de antioxidantrespons van het menselijk lichaam. Nrf2 is een eiwit dat niveaus van oxidatieve stress detecteert en de cellen in staat stelt zichzelf te beschermen tegen interne en externe factoren. Van Nrf2 is ook aangetoond dat het helpt bij het reguleren van genen die betrokken zijn bij de productie van antioxidante enzymen en stressresponsgenen. Het doel van het onderstaande artikel is om de effecten van Nrf2 bij kanker.
Abstract
De Keap1-Nrf2-route is de belangrijkste regulator van cytoprotectieve reacties op oxidatieve en elektrofiele stress. Hoewel celsignaleringsroutes die worden geactiveerd door de transcriptiefactor Nrf2 de initiatie en progressie van kanker in normale en premaligne weefsels voorkomen, biedt Nrf2-activiteit in volledig kwaadaardige cellen groeivoordeel door de chemoresistentie tegen kanker te vergroten en de groei van tumorcellen te verbeteren. In deze grafische review geven we een overzicht van de Keap1-Nrf2-route en de ontregeling ervan in kankercellen. We vatten ook kort de gevolgen samen van constitutieve Nrf2-activering in kankercellen en hoe dit kan worden benut bij kankergentherapie.
De Keap1-Nrf2-route is de belangrijkste regulator van cytoprotectieve reacties op endogene en exogene stress veroorzaakt door reactieve zuurstofsoorten (ROS) en elektrofielen [1]. De belangrijkste signaaleiwitten binnen de route zijn de transcriptiefactor Nrf2 (nucleaire factor erytroïde 2-gerelateerde factor 2) die samen met kleine Maf-eiwitten bindt aan het antioxidantresponselement (ARE) in de regulerende regio's van doelgenen, en Keap1 (Kelch ECH associërend eiwit 1), een repressoreiwit dat bindt aan Nrf2 en de afbraak ervan bevordert door de ubiquitine-proteasoomroute (Fig. 1). Keap1 is een zeer cysteïnerijk eiwit, muis Keap1 met in totaal 25 en menselijke 27 cysteïneresiduen, waarvan de meeste in vitro kunnen worden gemodificeerd door verschillende oxidanten en elektrofielen [2]. Van drie van deze residuen, C151, C273 en C288, is aangetoond dat ze een functionele rol spelen door de conformatie van Keap1 te veranderen, wat leidt tot nucleaire translocatie van Nrf2 en daaropvolgende doelgenexpressie [3] (Fig. 1). Het exacte mechanisme waardoor cysteïne-modificaties in Keap1 leiden tot Nrf2-activering is niet bekend, maar de twee overheersende maar niet wederzijds uitsluitende modellen zijn (1) het "scharnier en grendel"-model, waarin Keap1-modificaties in thiolresiduen die zich in de IVR van Keap1 bevinden verstoren de interactie met Nrf2 waardoor een verkeerde uitlijning van de lysine-residuen binnen Nrf2 wordt veroorzaakt die niet langer polyubiquitinyleerd kunnen worden en (2) het model waarin thiol-modificatie dissociatie van Cul3 van Keap1 veroorzaakt [3]. In beide modellen is de inductor-gemodificeerde en Nrf2-gebonden Keap1 geïnactiveerd en bijgevolg omzeilen nieuw gesynthetiseerde Nrf2-eiwitten Keap1 en verplaatsen zich naar de kern, binden aan de ARE en stimuleren de expressie van Nrf2-doelgenen zoals NAD (P) H chinon oxidoreductase 1 (NQO1), heem oxygenase 1 (HMOX1), glutamaat-cysteïne ligase (GCL) en glutathion S transferasen (GST's) (Fig. 2). Naast modificaties van Keap1-thiolen die resulteren in Nrf2-doelgeninductie, kunnen eiwitten zoals p21 en p62 binden aan Nrf2 of Keap1 en daardoor de interactie tussen Nrf2 en Keap1 verstoren [1], [3] (Fig. 3).
Fig. 1. Structuren van Nrf2 en Keap1 en de cysteïnecode. (A) Nrf2 bestaat uit 589 aminozuren en heeft zes evolutionair sterk geconserveerde domeinen, Neh1-6. Neh1 bevat een bZip-motief, een basisgebied - leucine-rits (L-Zip) -structuur, waarbij het basisgebied verantwoordelijk is voor DNA-herkenning en de L-Zip dimerisatie bemiddelt met kleine Maf-eiwitten. Neh6 functioneert als een degron om degradatie van Nrf2 in de kern te mediëren. Neh4 en 5 zijn transactiveringsdomeinen. Neh2 bevat ETGE- en DLG-motieven, die nodig zijn voor de interactie met Keap1 en een hydrofiel gebied van lysineresiduen (7 K), die onmisbaar zijn voor de Keap1-afhankelijke polyubiquitinatie en afbraak van Nrf2. (B) Keap1 bestaat uit 624 aminozuurresiduen en heeft vijf domeinen. De twee eiwit-eiwitinteractiemotieven, het BTB-domein en het Kelch-domein, worden gescheiden door het tussenliggende gebied (IVR). Het BTB-domein samen met het N-terminale deel van de IVR bemiddelt homodimerisatie van Keap1 en binding met Cullin3 (Cul3). Het Kelch-domein en het C-terminale gebied bemiddelen de interactie met Neh2. (C) Nrf2 interageert met twee moleculen van Keap1 via zijn Neh2 ETGE- en DLG-motieven. Zowel ETGE als DLG binden aan vergelijkbare plaatsen op het bodemoppervlak van het Keap1 Kelch-motief. (D) Keap1 is rijk aan cysteïneresiduen, met 27 cysteïnes in menselijk eiwit. Sommige van deze cysteïnes bevinden zich in de buurt van basische residuen en zijn daarom uitstekende doelwitten van elektrofielen en oxidanten. Het modificatiepatroon van de cysteïneresiduen door elektrofielen staat bekend als de cysteïnecode. De cysteïnecode-hypothese stelt voor dat structureel verschillende Nrf2-activerende middelen verschillende Keap1-cysteïnes beïnvloeden. De cysteïne-modificaties leiden tot conformationele veranderingen in de Keap1 die de interactie tussen de Nrf2 DLG- en Keap1 Kelch-domeinen verstoren, waardoor de polyubiquitinatie van Nrf2 wordt geremd. Het functionele belang van Cys151, Cys273 en Cys288 is aangetoond, aangezien Cys273 en Cys288 vereist zijn voor onderdrukking van Nrf2 en Cys151 voor activering van Nrf2 door inductoren [1], [3].
Fig. 2. De Nrf2-Keap1-signaleringsroute. (A en B) in basale omstandigheden binden twee Keap1-moleculen aan Nrf2 en Nrf2 wordt gepolyubiquityleerd door het op Cul3 gebaseerde E3-ligasecomplex. Deze polyubiquitilatie resulteert in snelle Nrf2-degradatie door het proteasoom. Een klein deel van Nrf2 ontsnapt aan het remmende complex en hoopt zich op in de kern om basale ARE-afhankelijke genexpressie te mediëren, waardoor de cellulaire homeostase behouden blijft. (C) Onder stressomstandigheden wijzigen inductoren de Keap1-cysteïnes die leiden tot de remming van Nrf2-ubiquitylering via dissociatie van het remmende complex. (D) Volgens het scharnier- en vergrendelingsmodel leidt modificatie van specifieke Keap1-cysteïneresiduen tot conformationele veranderingen in Keap1 wat resulteert in het losraken van het Nrf2 DLG-motief van Keap1. Ubiquitinatie van Nrf2 is verstoord, maar de binding met het ETGE-motief blijft. (E) In het Keap1-Cul3-dissociatiemodel wordt de binding van Keap1 en Cul3 verstoord als reactie op elektrofielen, wat leidt tot de ontsnapping van Nrf2 uit het alomtegenwoordige systeem. In beide voorgestelde modellen is de inductor-gemodificeerde en Nrf2-gebonden Keap1 geïnactiveerd en bijgevolg omzeilen nieuw gesynthetiseerde Nrf2-eiwitten Keap1 en verplaatsen zich naar de kern, binden aan het Antioxidant Response Element (ARE) en stimuleren de expressie van Nrf2-doelwit genen zoals NQO1, HMOX1, GCL en GST's [1], [3].
Fig. 3. Mechanismen voor constitutieve nucleaire accumulatie van Nrf2 bij kanker. (A) Somatische mutaties in Nrf2 of Keap1 verstoren de interactie van deze twee eiwitten. In Nrf2 beïnvloeden mutaties ETGE- en DLG-motieven, maar in Keap1 zijn mutaties gelijkmatiger verdeeld. Bovendien kan oncogenactivering, zoals KrasG12D [5], of verstoring van tumorsuppressoren, zoals PTEN [11] leiden tot transcriptionele inductie van Nrf2 en een toename van nucleair Nrf2. (B) Hypermethylering van de Keap1-promotor bij long- en prostaatkanker leidt tot vermindering van Keap1-mRNA-expressie, wat de nucleaire accumulatie van Nrf2 [6], [7] verhoogt. (C) Bij familiair papillair niercarcinoom leidt het verlies van fumaraathydratase-enzymactiviteit tot de accumulatie van fumaraat en verder tot succinatie van Keap1-cysteïneresiduen (2SC). Deze post-translationele modificatie leidt tot de verstoring van Keap1-Nrf2-interactie en nucleaire accumulatie van Nrf2 [8], [9]. (D) Accumulatie van disruptor-eiwitten zoals p62 en p21 kan de binding van Nrf2-Keap1 verstoren en resulteert in een toename van nucleair Nrf2. p62 bindt aan Keap1 en overlapt de bindingszak voor Nrf2 en p21 interageert direct met de DLG- en ETGE-motieven van Nrf2, waardoor het concurreert met Keap1 [10].
Mechanismen van activering en ontregeling van Nrf2 bij kanker
Hoewel cytoprotectie door Nrf2-activering belangrijk is voor chemopreventie van kanker in normale en premaligne weefsels, biedt Nrf2-activiteit in volledig kwaadaardige cellen groeivoordeel door de chemoresistentie tegen kanker te vergroten en de groei van tumorcellen te verbeteren [4]. Er zijn verschillende mechanismen beschreven waarmee de Nrf2-signaleringsroute constitutief wordt geactiveerd bij verschillende kankers: (1) somatische mutaties in Keap1 of het Keap1-bindingsdomein van Nrf2 die hun interactie verstoren; (2) epigenetische silencing van Keap1-expressie leidend tot defecte onderdrukking van Nrf2; (3) accumulatie van disruptor-eiwitten zoals p62, wat leidt tot dissociatie van het Keap1-Nrf2-complex; (4) transcriptionele inductie van Nrf2 door oncogene K-Ras, B-Raf en c-Myc; en (5) post-translationele modificatie van Keap1-cysteïnes door succinylatie die optreedt bij familiaal papillair niercarcinoom als gevolg van het verlies van fumaraathydratase-enzymactiviteit [3], [4], [5], [6], [7], [ 8], [9], [10] (afb. 3). Constitutief overvloedig Nrf2-eiwit veroorzaakt verhoogde expressie van genen die betrokken zijn bij het metabolisme van geneesmiddelen, waardoor de weerstand tegen chemotherapeutische geneesmiddelen en radiotherapie toeneemt. Bovendien is een hoog Nrf2-eiwitniveau geassocieerd met een slechte prognose bij kanker [4]. Overactief Nrf2 beïnvloedt ook de celproliferatie door glucose en glutamine naar anabole routes te sturen die de purinesynthese versterken en de pentosefosfaatroute beïnvloeden om celproliferatie te bevorderen [11] (Fig. 4).
Fig. 4. De dubbele rol van Nrf2 bij tumorigenese. Onder fysiologische omstandigheden zijn lage niveaus van nucleair Nrf2 voldoende voor het behoud van cellulaire homeostase. Nrf2 remt tumorinitiatie en kankermetastase door kankerverwekkende stoffen, ROS en andere DNA-beschadigende middelen te elimineren. Tijdens tumorigenese leidt accumulatie van DNA-schade tot constitutieve hyperactiviteit van Nrf2, wat de autonome kwaadaardige cellen helpt hoge niveaus van endogene ROS te verdragen en apoptose te voorkomen. Aanhoudend verhoogde nucleaire Nrf2-niveaus activeren metabole genen naast de cytoprotectieve genen die bijdragen aan metabole herprogrammering en verbeterde celproliferatie. Kankers met hoge Nrf2-niveaus worden geassocieerd met een slechte prognose vanwege radio- en chemoresistentie en agressieve proliferatie van kankercellen. De activiteit van de Nrf2-route is dus beschermend in de vroege stadia van tumorigenese, maar schadelijk in de latere stadia. Daarom blijft het verbeteren van de Nrf2-activiteit voor de preventie van kanker een belangrijke benadering, terwijl voor de behandeling van kanker Nrf2-remming wenselijk is [4], [11].
Aangezien hoge Nrf2-activiteit vaak voorkomt in kankercellen met nadelige resultaten, is er behoefte aan therapieën om Nrf2 te remmen. Helaas is de ontwikkeling van specifieke Nrf2-remmers vanwege structurele gelijkenis met sommige andere bZip-familieleden een uitdagende taak en zijn er tot nu toe slechts enkele onderzoeken naar Nrf2-remming gepubliceerd. Door natuurlijke producten te screenen, hebben Ren et al. [12] identificeerde een antineoplastische verbinding brusatol als een Nrf2-remmer die de chemotherapeutische werkzaamheid van cisplatine verbetert. Bovendien zijn PI3K-remmers [11], [13] en Nrf2-siRNA [14] gebruikt om Nrf2 in kankercellen te remmen. Onlangs hebben we een alternatieve benadering gebruikt, bekend als kanker-zelfmoordgentherapie, om kankercellen met hoge Nrf2-niveaus aan te pakken. Nrf2-aangedreven lentivirale vectoren [15] die thymidinekinase (TK) bevatten, worden overgebracht naar kankercellen met een hoge ARE-activiteit en de cellen worden behandeld met een pro-drug, ganciclovir (GCV). GCV wordt gemetaboliseerd tot GCV-monofosfaat, dat verder wordt gefosforyleerd door cellulaire kinasen tot een giftige trifosfaatvorm [16] (Fig. 5). Dit leidt tot effectieve doding van niet alleen TK-bevattende tumorcellen, maar ook van de aangrenzende cellen vanwege het omstandereffect [17]. ARE-gereguleerde TK/GCV-gentherapie kan verder worden verbeterd door een kankerchemotherapeutisch middel doxorubicine te combineren met de behandeling [16], wat het idee ondersteunt dat deze benadering nuttig zou kunnen zijn in combinatie met traditionele therapieën.
Fig. 5. Zelfmoordgentherapie. Constitutieve Nrf2-nucleaire accumulatie in kankercellen kan worden uitgebuit door Nrf2-aangedreven virale vector te gebruiken voor kanker-zelfmoordgentherapie [16]. In deze benadering wordt lentivirale vector (LV) die thymidinekinase (TK) tot expressie brengt onder minimale SV40-promoter met vier ARE's getransduceerd naar longadenocarcinoomcellen. Hoge nucleaire Nrf2-niveaus leiden tot robuuste expressie van TK via Nrf2-binding. Cellen worden vervolgens behandeld met een pro-drug, ganciclovir (GCV), die wordt gefosforyleerd door TK. Tri-gefosforyleerd GCV verstoort de DNA-synthese en leidt tot effectieve doding van niet alleen TK-bevattende tumorcellen, maar ook de naburige cellen vanwege het omstandereffect.
Nrf2 is een hoofdregulator die de productie van krachtige antioxidanten in het menselijk lichaam activeert die oxidatieve stress helpen elimineren. Verschillende antioxidante enzymen, zoals superoxide dismutase, of SOD, glutathion en catalase, worden ook geactiveerd via de Nrf2-route. Bovendien activeren bepaalde fytochemicaliën zoals kurkuma, ashwagandha, bacopa, groene thee en mariadistel Nrf2. Onderzoeksstudies hebben aangetoond dat Nrf2 activatie kan op natuurlijke wijze de cellulaire bescherming verbeteren en het evenwicht in het menselijk lichaam herstellen.
Dr. Alex Jimenez DC, CCST Insight
Sulforafaan en de effecten ervan op kanker, mortaliteit, veroudering, hersenen en gedrag, hartaandoeningen en meer
Isothiocyanaten zijn enkele van de belangrijkste plantaardige stoffen die u in uw dieet kunt krijgen. In deze video maak ik de meest uitgebreide case voor hen die ooit is gemaakt. Korte aandachtsspanne? Ga naar uw favoriete onderwerp door op een van de onderstaande tijdspunten te klikken. Volledige tijdlijn hieronder.
Belangrijkste secties:
00: 01: 14 - Kanker en sterfte
00: 19: 04 - Veroudering
00: 26: 30 - Hersenen en gedrag
00: 38: 06 - Laatste overzicht
00: 40: 27 - Dosis
Volledige tijdlijn:
00: 00: 34 - Introductie van sulforafaan, een belangrijk aandachtspunt van de video.
00: 01: 14 - Kruisbloemige groenteconsumptie en verlagingen van mortaliteit door alle oorzaken.
00: 02: 12 - Risico op prostaatkanker.
00: 02: 23 - Blaaskankerrisico.
00: 02: 34 - Longkanker bij rokers risico.
00: 02: 48 - Risico op borstkanker.
00: 03: 13 - Hypothetisch: wat als je al kanker hebt? (Interventie)
00: 03: 35 - Aannemelijk mechanisme dat de associatieve gegevens over kanker en mortaliteit stuurt.
00: 04: 38 - Sulforafaan en kanker.
00: 05: 32 - Dierlijk bewijs dat een sterk effect van extract van broccolispruit toont op de ontwikkeling van blaastumoren bij ratten.
00: 06: 06 - Effect van directe suppletie van sulforafaan bij prostaatkankerpatiënten.
00: 07: 09 - Bioaccumulatie van isothiocyanaatmetabolieten in daadwerkelijk borstweefsel.
00: 08: 32 - Remming van stamcellen van borstkanker.
00: 08: 53 - Geschiedenisles: brassica's werden vastgesteld met gezondheidseigenschappen zelfs in het oude Rome.
00: 09: 16 - het vermogen van Sulforaphane om de kankerverwekkende uitscheiding te verbeteren (benzeen, acroleïne).
00: 09: 51 - NRF2 als een genetische switch via antioxidantresponselementen.
00: 10: 10 - Hoe NRF2-activering de carcinogene excretie via glutathion-S-conjugaten verbetert.
00: 10: 34 - Spruitjes verhogen glutathion-S-transferase en verminderen DNA-schade.
00: 11: 20 - Broccolispruitendrank verhoogt de benzeenuitscheiding met 61%.
00: 13: 31 - Broccoli-spruithomogenaat verhoogt antioxiderende enzymen in de bovenste luchtwegen.
00: 15: 45 - Kruisbloemige groenteconsumptie en sterfte aan hart- en vaatziekten.
00: 16: 55 - Broccolispruitpoeder verbetert de bloedlipiden en het algemene risico op hartziekten bij type 2 diabetici.
00: 19: 04 - Begin van het verouderingsgedeelte.
00: 19: 21 - Met Sulforaphane verrijkte voeding verlengt de levensduur van kevers van 15 tot 30% (onder bepaalde omstandigheden).
00: 20: 34 - Belang van lage ontsteking voor een lang leven.
00: 22: 05 - Kruisbloemige groenten en broccoli-kiempoeder lijken een breed scala aan ontstekingsmarkers bij mensen te verminderen.
00: 24: 14 - Muisstudies suggereren dat sulforaphane de adaptieve immuunfunctie op oudere leeftijd zou kunnen verbeteren.
00: 25: 18 - Sulforafaan verbeterde de haargroei in een muismodel van kalend. Afbeelding bij 00: 26: 10.
00: 26: 30 - Begin van de sectie hersenen en gedrag.
00: 27: 18 - Effect van extract van broccolispruiten op autisme.
00: 27: 48 - Effect van glucoraphanine op schizofrenie.
00: 28: 17 - Begin van depressie discussie (plausibel mechanisme en studies).
00: 31: 21 - Muisstudie met behulp van 10 verschillende modellen van stress-geïnduceerde depressie laten sulforafaan zien, even effectief als fluoxetine (prozac).
00: 32: 00 - Onderzoek toont aan dat directe inname van glucoraphanine bij muizen even effectief is bij het voorkomen van depressies als gevolg van een sociaal nederlaagstressmodel.
00: 33: 01 - Begin van de sectie neurodegeneratie.
00: 33: 30 - Sulforafane en de ziekte van Alzheimer.
00: 33: 44 - Sulforafane en de ziekte van Parkinson.
00: 33: 51 - De ziekte van Sulforaphane en Hungtington.
00: 34: 43 - Begin van een traumatisch gedeelte over hersenletsel.
00: 35: 01 - Sulforafaan wordt geïnjecteerd onmiddellijk nadat TBI het geheugen verbetert (muisstudie).
00: 35: 55 - Sulforafaan en neuronale plasticiteit.
00: 36: 32 - Sulforafaan verbetert het leren in het model van type II diabetes bij muizen.
00: 37: 19 - Sulforafaan en duchenne spierdystrofie.
00: 37: 44 - Myostatinremming in spieratlanticellen (in vitro).
00: 38: 06 - Late video recapitulatie: sterfte en kanker, DNA-schade, oxidatieve stress en ontsteking, uitscheiding van benzeen, cardiovasculaire aandoeningen, type II diabetes, effecten op de hersenen (depressie, autisme, schizofrenie, neurodegeneratie), NRF2-route.
00: 40: 27 - Gedachten over het uitzoeken van een dosis broccoli of sulforafaan.
00: 41: 01 - Anekdotes over ontspruiten thuis.
00: 43: 14 - Op kooktemperaturen en sulforafaanactiviteit.
00: 43: 45 - Darmbacterieomzetting van sulforafaan uit glucoraphanine.
00: 44: 24 - Supplementen werken beter in combinatie met actieve myrosinase van groenten.
00: 44: 56 - Koken technieken en kruisbloemige groenten.
00: 46: 06 - Isothiocyanaten als goitrogens.
Dankwoord
Dit werk werd ondersteund door de Academie van Finland, de Sigrid Juselius Foundation en de Finse kankerorganisaties.
Concluderend, nucleaire factor (van erytroïde afgeleide 2)-achtige 2, ook bekend als NFE2L2 of Nrf2, is een eiwit dat de productie van antioxidanten verhoogt die het menselijk lichaam beschermen tegen oxidatieve stress. Zoals hierboven beschreven, wordt de stimulatie van de Nrf2-route onderzocht voor de behandeling van ziekten veroorzaakt door oxidatieve stress, waaronder kanker. De reikwijdte van onze informatie is beperkt tot chiropractische en spinale gezondheidsproblemen. Als u het onderwerp wilt bespreken, kunt u het aan Dr. Jimenez vragen of contact met ons opnemen via�915-850-0900 .
Extra topic discussie: Kniepijn verlichten zonder operatie
Kniepijn is een bekend symptoom dat kan optreden als gevolg van een verscheidenheid aan knieblessures en / of aandoeningen, waaronder sportverwondingen. De knie is een van de meest complexe gewrichten in het menselijk lichaam omdat deze bestaat uit de kruising van vier botten, vier ligamenten, verschillende pezen, twee menisci en kraakbeen. Volgens de American Academy of Family Physicians zijn de meest voorkomende oorzaken van kniepijn onder patellaire subluxatie, patella tendinitis of jumper's knee en Osgood-Schlatter-ziekte. Hoewel kniepijn het meest voorkomt bij mensen ouder dan 60 jaar, kan kniepijn ook voorkomen bij kinderen en adolescenten. Kniepijn kan thuis worden behandeld volgens de RICE-methoden; ernstige knieverwondingen kunnen echter onmiddellijke medische aandacht vereisen, inclusief chiropractische zorg.
DNA ondersteunt ongeveer 20,000-genen, elk met een programma voor het creëren van een eiwit of enzym dat nodig is voor een gezonde levensstijl. Elk van deze patronen moet constant worden gereguleerd door een soort van 'promotor' die precies weet hoeveel van elke stof en / of chemische stof wordt gegenereerd en onder welke omstandigheden deze zich ook zullen ontwikkelen.
Door verbinding te maken met een bepaald soort switch-achtige promotorgebieden, bekend als het Antioxidant Response Element, of ARE, de Nrf2-factor�ondersteunt de aanmaaksnelheid van honderden verschillende genen die de cellen in staat stellen te overleven onder stressvolle omstandigheden. Deze genen genereren vervolgens een selectie van antioxidante enzymen die een verdedigingsnetwerk ontwikkelen door oxidanten te neutraliseren en door de giftige bijproducten die bij hun productie achterblijven op te ruimen, naast het helpen herstellen van de schade die ze hebben veroorzaakt.
Wat is oxidatieve stress?
Verschillende oxidatiemiddelen, zoals het superoxide-radicaal, of O2-., en waterstofperoxide, of H2O2, zijn gecreëerd door het verbranden van de stoffen en/of chemicaliën die het menselijk lichaam ondersteunen. Het menselijk lichaam bezit antioxiderende enzymen die reactieve voedingsmiddelen en dranken die we consumeren neutraliseren en ontgiften. De Nrf2 moduleert hun productie om het evenwicht te bewaren en onderstreept de vraag naar al deze enzymen. Dit evenwicht kan worden verstoord door een aantal factoren, waaronder leeftijd.
Naarmate we ouder worden, creëert het menselijk lichaam minder Nrf2 en dit delicate evenwicht kan geleidelijk beginnen te draaien naar de oxidatieve kant, een toestand die oxidatieve stress wordt genoemd. Ziekte kan ook de overproductie van oxidanten veroorzaken. Infecties, allergieën en auto-immuunziekten kunnen onze immuuncellen bovendien aanzetten tot het aanmaken van reactieve oxidanten, zoals O2-. , H2O2, OH en HOCl, waar gezonde cellen beschadigd raken en reageren met ontstekingen. Ziekten die verband houden met veroudering, waaronder hartaanvallen, beroertes, kanker en neurodegeneratieve aandoeningen zoals de ziekte van Alzheimer, verhogen ook de ontwikkeling van oxidanten, waardoor stress en een ontstekingsreactie ontstaan.
Wat zijn Nrf2-activators?
Het Nrf2-eiwit, ook wel een transcriptiefactor genoemd vanwege de manier waarop het enzymen en genen kan ondersteunen en regelen, is het geheime element van een reeks biochemische reacties in de cel die reageert op modificaties in cognitief evenwicht en oxidatieve balans. De sensorelementen van dit pad wijzigen en ontladen Nrf2, waardoor het wordt geactiveerd, zodat het zich in de kern van de cel naar het DNA kan verspreiden. De Nrf2 kan als alternatief de genen en enzymen die het ondersteunt inschakelen of uitschakelen om de cel te beschermen.
Gelukkig is er een verscheidenheid aan stoffen die Nrf2-activators ontwikkelen door het verbruik van bepaalde planten en extracten die eeuwen geleden in Chinese en Indiaanse traditionele remedies werden gebruikt. Deze fytochemicaliën lijken even krachtig te zijn met minder bijwerkingen, als de Nrf2-activerende farmaceutische producten die tegenwoordig worden gebruikt.
Nucleaire factor erytroïde 2-gerelateerde factor, beter bekend als Nrf2, is een transcriptiefactor die de cel beschermt door genen, enzymen en antioxidantreacties te reguleren. Transcriptiefactoren zijn een soort eiwit die zich hechten aan DNA om de aanmaak van specifieke stoffen en chemicaliën te bevorderen, waaronder glutathione S-transferasen of GST's. Nrf2-activatie induceert de productie van actieve eiwitten die een krachtig antioxidantvermogen vertonen om oxidatieve stress te verminderen.
Dr. Alex Jimenez DC, CCST Insight
De wetenschap achter Nrf2-activering
Nadat het oorspronkelijke Nrf2-activerende voedingssupplement in 2004 was aangemaakt, was er minimale informatie bekend over de functie van de Nrf2-route. Ongeveer 200 kranten in de literatuur over Nrf2, ook bekend als nucleair factor-achtige 2 of NFE2L2, bestonden en onderzoekers waren nog maar net begonnen met het ontdekken van de antioxidant reactie van Nrf2 bij zoogdieren. Vanaf 2017 zijn echter via 9,300 academische onderzoeksstudies over deze "hoofdregelaar" afgedrukt.
In werkelijkheid regelt Nrf2 veel antioxidante enzymen die niet correleren met de genen, maar bieden ze bescherming tegen een verscheidenheid aan stressgerelateerde omstandigheden die worden aangetroffen door cellen, organen en uiteindelijk organismen, onder gezonde en pathologische omstandigheden. Op basis van deze nieuwe hoeveelheid informatie uit gepubliceerde wetenschappelijke onderzoeken, kunnen onderzoekers zich nu beter ontwikkelen Nrf2 voedingssupplementen.
Vanaf 2007 hebben onderzoeksstudies de complexe functie van de Nrf2-route aangetoond. Er is gevonden dat Nrf2-activators factoren van verschillende structuren in het menselijk lichaam nabootsen. Via deze routes zijn Nrf2-activators uitgerust om veranderende omstandigheden in de cel te voelen om het evenwicht te bewaren en te reageren op de veranderende vereisten van de genen.
Waarom Nrf2 gebruiken-Supplementen activeren?
Omdat de capaciteiten van Nrf2-activering afnemen met de leeftijd in organismen, kunnen er veranderingen optreden. Onderzoeksstudies hebben aangetoond dat de focus van Nrf2 in cellen afneemt met de leeftijd, wat leidt tot verhoogde markers van oxidatieve stress. Een verscheidenheid aan leeftijdsgerelateerde ziekten zoals atherosclerose en cardiovasculaire aandoeningen, artritis, kanker, obesitas, type 2 diabetes, hypertensie, cataract en de ziekte van Alzheimer en Parkinson kunnen zich ontwikkelen als gevolg van deze veranderingen. Oxidatieve stress is gevonden met deze gezondheidsproblemen.
Door het vermogen van de cel om de productie van Nrf2-activatoren te verhogen te stimuleren, Nrf2 voedingssupplementen kan helpen om het vermogen van het menselijk lichaam om de effecten van oxidatieve stress tegen te gaan, te doen herleven. Meervoudig onverzadigde vetzuren, of PUFA's, zijn een van de gemakkelijkst geoxideerde moleculen en ze zijn bijzonder kwetsbaar voor schade door vrije radicalen. De productie van thiobarbituurzuur of TBARS kan met de leeftijd toenemen, wat wijst op verhoogde oxidatieve stress en een daling in Nrf2-gereguleerde pathways.
Biologisch gezien is gen-inductie een heel langzaam mechanisme, dat over het algemeen uren nodig heeft om via een route over te gaan. Als gevolg hiervan hebben veel enzymen hun eigen aan/uit-schakelaars die binnen enkele minuten kunnen worden geactiveerd door verschillende regulerende enzymen. Onderzoekers hebben eigen composities van Nrf2-activators ontwikkeld die gebruikmaken van deze kennisbank van activering. Nrf2-activering bestaat niet alleen uit de Nrf2-transcriptiefactor die wordt ontladen uit zijn remmer en migreert naar de celkern, maar ook door binding aan specifieke DNA-sequenties om cytoprotectieve genexpressie aan te moedigen, waardoor het tempo wordt gereguleerd waarmee Nrf2 uit de kern wordt gehaald.
Door de eliminatieprocedure en de activering van Nrf2 in het menselijk lichaam te begrijpen, hebben onderzoekers combinaties van verschillende Nrf2-activators kunnen bouwen om de reflectie van genen te bereiken door middel van modulatie. De combinatie van de kennisbank, samen met de grote verscheidenheid aan andere onderzoeksstudies, heeft geholpen om Nrf2-activators te produceren voor gebruik als voedingssupplementen. De reikwijdte van onze informatie is beperkt tot chiropractische en spinale gezondheidsproblemen. Als u het onderwerp wilt bespreken, kunt u het aan Dr. Jimenez vragen of contact met ons opnemen via�915-850-0900 .
Curator van Dr. Alex Jimenez
Extra topic discussie: Kniepijn verlichten zonder operatie
Kniepijn is een bekend symptoom dat kan optreden als gevolg van een verscheidenheid aan knieblessures en / of aandoeningen, waaronder sportverwondingen. De knie is een van de meest complexe gewrichten in het menselijk lichaam omdat deze bestaat uit de kruising van vier botten, vier ligamenten, verschillende pezen, twee menisci en kraakbeen. Volgens de American Academy of Family Physicians zijn de meest voorkomende oorzaken van kniepijn onder patellaire subluxatie, patella tendinitis of jumper's knee en Osgood-Schlatter-ziekte. Hoewel kniepijn het meest voorkomt bij mensen ouder dan 60 jaar, kan kniepijn ook voorkomen bij kinderen en adolescenten. Kniepijn kan thuis worden behandeld volgens de RICE-methoden; ernstige knieverwondingen kunnen echter onmiddellijke medische aandacht vereisen, inclusief chiropractische zorg.
Antioxidanten worden wetenschappelijk aangeduid als verbindingen die het oxidatieproces in het menselijk lichaam beperken, en als ze niet worden gecontroleerd, kunnen er vrije radicalen ontstaan die tal van kettingreacties kunnen ontwikkelen die cellulaire schade kunnen veroorzaken. Gelukkig kan het menselijk lichaam dergelijke ingebouwde immuunmechanismen creëren, maar wanneer het opzetten van reactieve zuurstofspecies, of ROS, niet in staat is om te worden geneutraliseerd, stel je een klein vlammetje voor dat uit de hand loopt wanneer het wordt doordrenkt met zuurstof, zal er zeker schade optreden. .
Om verder te gaan met de metafoor van de vlam: het uiteindelijke product van het niet kunnen neutraliseren van de impact van ROS, of reactieve zuurstofsoorten, is zowel schade als ontsteking, met andere woorden, het menselijk lichaam staat letterlijk in brand. Het fantastische is dat er antioxidanten zijn die enorm kunnen helpen bij het bestrijden van dit gezondheidsprobleem en deze antioxidant is glutathion. Hoewel gevonden in 1889, is het antioxiderende effect van glutathion een van de meest interessante onderwerpen geworden in moderne onderzoeken.
Meester van antioxidanten: glutathion
De krachtige stof is een tripeptide dat ontstaat uit cysteïne, glutaminezuur en glycine. Vanwege zijn vermogen om het menselijk lichaam te beschermen tegen het ontstaan van vrije radicalen, kan glutathion uiteindelijk helpen bij het bevorderen van een gezond immuunsysteem. Gebaseerd op Wetenschappelijke rapporten in 2015, werd vastgesteld dat het vermogen van glutathion om synergistisch te functioneren met peroxiredine en katalase cellen helpt beschermen tegen waterstofperoxide. Deze synergetische formule werkt tegen reactieve zuurstofspecies of ROS. Glutathion, peroxidredine en catalase zijn essentiële elementen in de toename van cellulaire homeostase, wat een essentieel proces is van gezonde cellen, weefsels en organen.
Bovendien verhoogt glutathion de algehele structuur en functie van het immuunsysteem door gebruik te maken van het belangrijke effect ervan op de lymfocytenfuncties. Volgens de Afdeling Immunochemie, kan het correct aanvullen van niveaus van glutathion in het menselijk lichaam de immuunreacties aanzienlijk verbeteren. Twee gerandomiseerde, placebogecontroleerde onderzoeken toonden bijvoorbeeld aan dat de therapeutische behandeling van immuungecompromitteerde patiënten met N-acetylcysteïne, of NAC, in beide gevallen resulteerde in een substantiële groei van de meeste immunologische processen, waaronder een volledige verjonging activiteit van natuurlijke killercellen. N-acetylcysteïne, of NAC, gebruikt de zwavel uit glutathion en combineert deze met giftige moleculen, die vervolgens in water oplosbaar worden en in het menselijk lichaam worden uitgescheiden.
Glutathion heeft ook het vermogen om liponzuur nieuw leven in te blazen en vitamine C en E te recyclen, die nodig zijn om bepaalde systeemprocessen op gang te brengen door elektronen te sturen om vrije radicalen te neutraliseren. Gebaseerd op een onderzoek van PLoS ONE, glutathion beïnvloedde patiënten met diabetes metillus, of T2DM, en mycobacterium tuberculosis. Normaal gesproken hebben personen met een zwak immuunsysteem de neiging om meer blootgesteld te worden aan M. tb, of mycobacterium tuberculosis, ziekte of infectie. Bovendien zijn personen met diabetes metilllus type 2, of T2DM, twee tot drie keer meer vatbaar voor tuberculose dan mensen zonder T2DM. De onderzoeksstudie suggereerde ook dat het verhogen van de niveaus van glutathion in macrofagen geïsoleerd van patiënten met T2DM leidde tot een betere controle van M.Tb-ziekte of -infectie. Deze resultaten tonen aan dat lagere niveaus van glutathion bij patiënten met T2DM bijdragen aan een verhoogde kans op de ziekte of infectie van M. tb. Bovendien afhankelijk van Dietro Ghezzi in Brighton en Sussex Medical School, kan oxidatieve stress uiteindelijk leiden tot een slechte structuur en functie van het immuunsysteem.
Gelukkig speelt glutathion een essentiële rol bij het versterken en beheersen van de immuniteit. Glutathion is bijvoorbeeld essentieel voor aangeboren en adaptieve processen binnen het immuunsysteem, waaronder proliferatie van T-lymfocyten, fagocytische activiteit van polymorfonucleaire neutrofielen en dendritische celfuncties, die van fundamenteel belang kunnen zijn omdat deze bestaan uit antigeenpresenterende cellen. . Celgemedieerde immuniteit omvat eiwitantigenen die aanvankelijk beginnen te degenereren in de endocytische blaasjes van macrofagen en dendritische cellen, daarom wordt aangetoond dat de kleinere peptiden op het oppervlak de proliferatie van antigeenspecifieke T-cellen activeren. Bovendien helpt glutathion bij de aanmaak van cytokines en is het noodzakelijk om de productie van interferon-gamma door dendritische cellen in stand te houden, wat belangrijk is voor de bescherming tegen intracellulaire pathogenen, waaronder mycobacteriën.
N-acetyl-cysteïne, of NAC, wetenschappelijk aangeduid als de voorloper van glutathion, is ook een zeer krachtige cellulaire antioxidant die wordt gebruikt als antioxidant voor het wegvangen van vrije radicalen. Algemeen erkend voor zijn rol in acetaminophen-toxiciteit voorkomenVan NAC, of N-acetyl-cysteïne, is aangetoond dat het verschillende gezondheids- en welzijnsvoordelen bezit. Volgens celjournaal, helpt NAC een gezonde ontstekingsreactie te ondersteunen en kan het een positieve invloed hebben op de menselijke termijn en vroeggeboorte. De onderzoeksstudie concludeerde dat bij vrouwen met eerdere vroeggeboorte en bacteriële vaginose, 0.6 gram NAC per dag oraal ingenomen samen met progesteron na week 16 van de zwangerschap beschermd tegen vroeggeboorte herhaling en verbeterde neonatale uitkomst. Concluderend werden ook de positieve effecten van NAC op de spieropbouw gedetecteerd. Na drie minuten van aanhoudende samentrekkingen was er een 15 procent verbeterde output, wat aantoont hoe NAC een fundamentele rol speelt bij het verbeteren van spieropbouw en het verminderen van algehele vermoeidheid tijdens de bevalling.
Onderzoekers ontdekten ook dat NAC, of N-acetyl-cysteïne, gunstig kan zijn voor mensen met polycysteus ovariumsyndroom of PCOS. PCOS, of polycysteus ovariumsyndroom, is een veel voorkomende endocriene klierziekte die ongeveer 5 tot 10 procent van de vrouwen in de vruchtbare leeftijd treft. Bij dergelijke patiënten is er een groter risico op het ervaren van het metabool syndroom, waarbij het gebruik van NAC hielp bij het herstellen van gezonde insulinespiegels en gevoeligheid.
Insight van Dr. Alex Jimenez
Glutathion wordt de "meester van antioxidanten" genoemd vanwege zijn fundamentele rol bij het bereiken en behouden van algehele gezondheid en welzijn. Hoewel het menselijk lichaam in staat is om zijn eigen glutathion te produceren, kunnen slechte voeding, vervuiling, gifstoffen, overmatig gebruik van drugs en/of medicijnen, stress, trauma, veroudering, ziekte en straling allemaal onze natuurlijke niveaus van glutathion verlagen. Dit kan op zijn beurt individuen vatbaarder maken voor celbeschadiging door oxidatieve stress, vrije radicalen, infecties en kanker. Suppletie met glutathion kan daarom enorme voordelen hebben voor het menselijk lichaam. Samen met alternatieve behandelingsopties, zoals chiropractische zorg, kunnen glutathionspiegels opnieuw worden gereguleerd om het welzijn te verbeteren.
Bovendien hebben beroepsbeoefenaren in de gezondheidszorg voorgesteld om het gebruik van glutathion-suppletie te implementeren in combinatie met andere alternatieve behandelingsopties, zoals chiropractische zorg, om de algehele gezondheid en het welzijn verder te verbeteren. Antioxidanten zijn belangrijk voor het behoud van maximaal welzijn en om de kettingreactie van vrije radicalen die celbeschadiging of -beschadiging veroorzaken te remmen. Krachtige antioxidanten zoals glutathion helpen, zoals eerder vermeld, uiteindelijk bij het reguleren van de ontwikkeling van deze vrije radicalen en zorgen voor een gezondere reactie van het immuunsysteem. Onderzoeksstudies hebben dat uitgewezen chiropractische zorg kan ook een essentiële rol spelen in dit proces, waarbij de activiteit van antioxidanten in het menselijk lichaam op natuurlijke wijze wordt gestimuleerd. Chiropractische zorg is een veilige en effectieve behandelmethode waarbij gebruik wordt gemaakt van spinale aanpassingen en manuele manipulaties om afwijkingen van de wervelkolom of subluxaties te corrigeren, zodat het menselijk lichaam zichzelf op natuurlijke wijze kan genezen zonder het gebruik van medicijnen/medicijnen en/of chirurgische ingrepen.
Ten slotte demonstreren antioxidanten hun biologische eigenschappen door middel van een groot aantal gezondheidsvoordelen, die nu meer dan ooit nodig kunnen zijn met de steeds toenemende aanval van stress, ziekte en vervuiling in onze moderne wereld, die allemaal bijdragen aan celbeschadiging en/of -beschadiging . Glutathion en zijn voorloper, NAC, of N-acetyl-cysteïne, blijven hun krachtige status op het gebied van antioxidanten tonen. Samen met alternatieve behandelingsopties, zoals chiropractische zorg, kunnen mensen profiteren van alle voordelen die deze krachtige antioxidant te bieden heeft. De reikwijdte van onze informatie is beperkt tot chiropractie en ruggenmergletsels en -aandoeningen. Om het onderwerp te bespreken, kunt u het Dr. Jimenez vragen of contact met ons opnemen via�915-850-0900 .
Curator van Dr. Alex Jimenez
Bijkomende onderwerpen: Rugpijn
Rugpijn is een van de meest voorkomende oorzaken van invaliditeit en gemiste dagen op het werk wereldwijd. Rugpijn is trouwens toegeschreven als de op één na meest voorkomende reden voor bezoeken aan artsenbureaus, die in de minderheid zijn dan alleen infecties aan de bovenste luchtwegen. Ongeveer 80 procent van de bevolking ervaart ten minste één keer gedurende hun hele leven een vorm van rugpijn. De wervelkolom is een complexe structuur bestaande uit botten, gewrichten, ligamenten en spieren, naast andere zachte weefsels. Vanwege dit, verwondingen en / of verergerde omstandigheden, zoals herniated discs, kan uiteindelijk leiden tot symptomen van rugpijn. Sportblessures of ongevallen met auto-ongelukken zijn vaak de meest voorkomende oorzaak van rugpijn, maar soms kunnen de eenvoudigste bewegingen pijnlijke resultaten hebben. Gelukkig kunnen alternatieve behandelingsopties, zoals chiropractische zorg, de rugpijn verlichten door het gebruik van spinale aanpassingen en handmatige manipulaties, waardoor uiteindelijk de pijnverlichting wordt verbeterd.
Science based Chiropractor Dr. Alexander Jimenez neemt een kijkje op oxidatieve stress, wat het is, hoe het het lichaam beïnvloedt en de antioxidantverdediging om de situatie te verhelpen.
Esra Birben PhD, 1 Umit Murat Sahiner MD, 1 Cansin Sackes MD, 1 Serpil Erzurum MD, 2 en Omer Kalayci, MD1
Samenvatting: Reactieve zuurstofspecies (ROS) worden geproduceerd door levende organismen als gevolg van een normaal cellulair metabolisme en omgevingsfactoren, zoals luchtverontreinigende stoffen of sigarettenrook. ROS zijn zeer reactieve moleculen en kunnen celstructuren zoals koolhydraten, nucleïnezuren, lipiden en eiwitten beschadigen en hun functies veranderen. De verschuiving in de balans tussen oxidanten en antioxidanten ten gunste van oxidanten wordt 'oxidatieve stress' genoemd. Regulatie van de reducerende en oxiderende (redox) toestand is cruciaal voor de levensvatbaarheid, activering, proliferatie en orgaanfunctie van de cel. Aërobe organismen hebben geïntegreerde antioxidantsystemen, waaronder enzymatische en niet-enzymatische antioxidanten die meestal effectief zijn in het blokkeren van schadelijke effecten van ROS. In pathologische omstandigheden kunnen de antioxidantsystemen echter overweldigd worden. Oxidatieve stress draagt bij aan vele pathologische aandoeningen en ziekten, waaronder kanker, neurologische aandoeningen, atherosclerose, hypertensie, ischemie/perfusie, diabetes, acute respiratory distress syndrome, idiopathische longfibrose, chronische obstructieve longziekte en astma. In deze review vatten we de cellulaire oxidant- en antioxidantsystemen samen en bespreken we de cellulaire effecten en mechanismen van de oxidatieve stress.
Reactieve zuurstofspecies (ROS) worden geproduceerd door levende organismen als gevolg van normaal cellulair metabolisme. Bij lage tot matige concentraties functioneren ze in fysiologische celprocessen, maar bij hoge concentraties veroorzaken ze nadelige modificaties aan celcomponenten, zoals lipiden, eiwitten en DNA.1 De verschuiving in de balans tussen oxidant/antioxidant ten gunste van oxidanten wordt 'oxidatieve stress' genoemd. Oxidatieve stress draagt bij aan vele pathologische aandoeningen, waaronder kanker, neurologische aandoeningen, 6 atherosclerose, hypertensie, ischemie/perfusie, 7 diabetes, acute respiratory distress syndrome, idiopathische longfibrose, chronische obstructieve longziekte ,10 en astma.11�14 Aërobe organismen hebben geïntegreerde antioxidantsystemen, waaronder enzymatische en niet-enzymatische antioxidanten die gewoonlijk effectief zijn in het blokkeren van de schadelijke effecten van ROS. In pathologische omstandigheden kunnen de antioxidantsystemen echter overweldigd worden. In deze review vatten we de cellulaire oxidant- en antioxidantsystemen en regulering van de reducerende en oxiderende (redox) toestand in gezondheids- en ziektetoestanden samen.
oxidanten
Endogene bronnen van ROS
ROS worden geproduceerd uit moleculaire zuurstof als een resultaat van normaal cellulair metabolisme. ROS kan worden onderverdeeld in 2-groepen: vrije radicalen en niet-radicalen. Moleculen die één of meer ongepaarde elektronen bevatten en aldus reactiviteit aan het molecuul geven, worden vrije radicalen genoemd. Wanneer 2-vrije radicalen hun ongepaarde elektronen delen, worden niet-radicale vormen gemaakt. De belangrijkste ROS van 3 die van fysiologische betekenis zijn, zijn superoxide-anion (O22.), Hydroxylradicaal (OH) en waterstofperoxide (H2O2). ROS zijn samengevat in tabel 1.
Superoxide-anion wordt gevormd door de toevoeging van 1 elektron aan de moleculaire zuurstof.22 Dit proces wordt gemedieerd door nicotine-adenine-dinucleotidefosfaat [NAD(P)H]-oxidase of xanthine-oxidase of door het mitochondriale elektronentransportsysteem. De belangrijkste plaats voor de productie van superoxide-anionen zijn de mitochondriën, de machinerie van de cel om adenosinetrifosfaat te produceren. Normaal gesproken worden elektronen overgedragen via de mitochondriale elektronentransportketen voor reductie van zuurstof tot water, maar ongeveer 1 tot 3% van alle elektronen lekt uit het systeem en produceert superoxide. NAD(P)H-oxidase wordt aangetroffen in polymorfonucleaire leukocyten, monocyten en macrofagen. Bij fagocytose produceren deze cellen een uitbarsting van superoxide die leidt tot bacteriedodende activiteit. Superoxide wordt omgezet in waterstofperoxide door de werking van superoxidedismutasen (SOD's, EC 1.15.1.1). Waterstofperoxide diffundeert gemakkelijk door het plasmamembraan. Waterstofperoxide wordt ook geproduceerd door xanthine-oxidase, aminozuuroxidase en NAD(P)H-oxidase�23,24 en in peroxisomen door consumptie van moleculaire zuurstof bij metabolische reacties. In een opeenvolging van reacties die Haber�Weiss- en Fenton-reacties worden genoemd, kan H2O2 worden afgebroken tot OH2 in aanwezigheid van transmissiemetalen zoals Fe21 of Cu21.25
Fe31 +�.O2�?Fe2 +�O2 Haber Weiss
Fe2 +�H2O2�?Fe3 +�OH�+ .OH Fenton-reactie
O 2 �zelf kan ook reageren met H2 O2 en OH genereren�.26,27 Hydroxylradicaal is het meest reactieve van ROS en kan eiwitten, lipiden, koolhydraten en DNA beschadigen. Het kan ook lipideperoxidatie starten door een elektron uit meervoudig onverzadigde vetzuren te nemen.
Granulocytische enzymen breiden de reactiviteit van H2O2 verder uit via eosinofielperoxidase en myeloperoxidase (MPO). In geactiveerde neutrofielen wordt H2O2 verbruikt door MPO. In aanwezigheid van chloride-ionen wordt H2O2 omgezet in hypochloorzuur (HOCl). HOCl is zeer oxidatief en speelt een belangrijke rol bij het doden van de ziekteverwekkers in de luchtwegen.28 HOCl kan echter ook reageren met DNA en DNA-eiwitinteracties induceren en pyrimidine-oxidatieproducten produceren en chloride toevoegen aan DNA-basen.29,30 Eosinophil peroxidase en MPO dragen ook bij aan de oxidatieve stress door modificatie van eiwitten door halogeneringen, nitratie en eiwitverknopingen via tyrosylradicalen.31
Andere van zuurstof afgeleide vrije radicalen zijn de peroxylradicalen (ROO $). De eenvoudigste vorm van deze radicalen is hydroperoxylradicaal (HOO $) en speelt een rol bij de peroxidatie van vetzuren. Vrije radicalen kunnen lipideperoxidatie-kettingreacties veroorzaken door een waterstofatoom te abstraheren uit een methyleenkoolstof met zijketens. De lipidische rest reageert dan met zuurstof om peroxylradicaal te produceren. Peroxylradicaal initieert een kettingreactie en transformeert meervoudig onverzadigde vetzuren in lipide hydroperoxiden. Lipidehydroperoxiden zijn zeer onstabiel en kunnen gemakkelijk worden afgebroken tot secundaire producten, zoals aldehyden (zoals 4-hydroxy-2,3-nonenal) en malondialdehyden (MDA's). Isoprostanen zijn een andere groep lipideperoxidatieproducten die worden gegenereerd via de peroxidatie van arachidonzuur en die ook zijn verhoogd in plasma- en ademcondensaten van astmatici. 34,35 Peroxidatie van lipiden verstoort de integriteit van celmembranen en leidt tot herschikking van de membraanstructuur .
Waterstofperoxide, superoxide-radicaal, geoxideerd glutathion (GSSG), MDA's, isoprostanen, carbonylen en nitrotyrosine kunnen eenvoudig worden gemeten uit plasma-, bloed- of bronchoalveolaire lavage-monsters als biomarkers voor oxidatie door gestandaardiseerde assays.
Exogene bron van oxidanten
Sigarettenrook
Sigarettenrook bevat veel oxidanten en vrije radicalen en organische verbindingen, zoals superoxide en stikstofoxide.36 Daarnaast activeert inhalatie van sigarettenrook in de longen ook enkele endogene mechanismen, zoals ophoping van neutrofielen en macrofagen, die de oxidantbeschadiging verder vergroten .
Ozonblootstelling
Ozonblootstelling kan lipideperoxidatie veroorzaken en de instroom van neutrofielen in het epitheel van de luchtwegen induceren. Kortdurende blootstelling aan ozon veroorzaakt ook de afgifte van ontstekingsmediatoren, zoals MPO, eosinofiele kationische eiwitten en ook lactaatdehydrogenase en albumine. 37 Zelfs bij gezonde proefpersonen veroorzaakt blootstelling aan ozon een vermindering van de longfuncties.38 Cho et al39 hebben aangetoond dat fijn stof (mengsel van vaste deeltjes en vloeibare druppels gesuspendeerd in de lucht) katalyseert de reductie van zuurstof.
hyperoxie
Hyperoxia verwijst naar aandoeningen met hogere zuurstofniveaus dan de normale partiële zuurstofdruk in de longen of andere lichaamsweefsels. Het leidt tot een grotere productie van reactieve zuurstof- en stikstofsoorten. 40,41
Ioniserende straling
Ioniserende straling zet in aanwezigheid van O2 hydroxylradicaal, superoxide en organische radicalen om in waterstofperoxide en organische hydroperoxiden. Deze hydroperoxidesoorten reageren met redox-actieve metaalionen, zoals Fe en Cu, via Fenton-reacties en induceren zo oxidatieve stress.42,43 Narayanan et al44 toonden aan dat fibroblasten die werden blootgesteld aan alfadeeltjes een significante toename van intracellulaire O2 2. en H2O2 hadden. productie via plasmamembraangebonden NADPH-oxidase.44 Signaaltransductiemoleculen, zoals extracellulaire signaalgereguleerde kinase 1 en 2 (ERK1/2), c-Jun N-terminale kinase (JNK) en p38, en transcriptiefactoren, zoals activator eiwit-1 (AP-1), nucleaire factor-kB (NF-kB) en p53 worden geactiveerd, wat resulteert in de expressie van stralingsresponsgerelateerde genen.45 Ultraviolet A (UVA) fotonen veroorzaken oxidatieve reacties door excitatie van endogene fotosensibilisatoren, zoals porfyrines, NADPH-oxidase en riboflavines. 50-Oxo-8-dihydroguanine (7,8-oxoGua) is het belangrijkste UVA-gemedieerde DNA-oxidatieproduct dat wordt gevormd door de oxidatie van OH-radicaal, 8-elektron-oxidanten en singlet-zuurstof dat voornamelijk reageert met guanine.1 De vorming van guanine Het is aangetoond dat radicale kationen in geïsoleerd DNA efficiënt optreden door het directe effect van ioniserende straling.51 Na blootstelling aan ioniserende straling neemt het intracellulaire niveau van glutathion (GSH) gedurende korte tijd af, maar neemt daarna weer toe.52,53
Heavy Metal Ionen
Zwaar metaalionen, zoals ijzer, koper, cadmium, kwik, nikkel, lood en arseen, kunnen de vorming van reactieve radicalen veroorzaken en cellulaire schade veroorzaken door uitputting van enzymactiviteiten door lipideperoxidatie en reactie met nucleaire eiwitten en DNA.55
Een van de belangrijkste mechanismen van metaalgemedieerde vorming van vrije radicalen is via een Fenton-type reactie. Superoxide-ionen en waterstofperoxide kunnen interageren met overgangsmetalen, zoals ijzer en koper, via de door metaal gekatalyseerde Haber-Weiss/Fenton-reactie om OH-radicalen te vormen.
Naast de mechanismen van het Fenton-type en het Haber-Weiss-type, kunnen bepaalde metaalionen direct reageren met cellulaire moleculen om vrije radicalen te genereren, zoals thiolradicalen, of celsignaleringsroutes te induceren. Deze radicalen kunnen ook reageren met andere thiolmoleculen om O22.. O22 te genereren. wordt omgezet in H2O2, wat zorgt voor extra vorming van zuurstofradicalen. Sommige metalen, zoals arseniet, induceren ROS-vorming indirect door activering van radicaalproducerende systemen in cellen.56
Arseen is een zeer giftig element dat een verscheidenheid aan ROS produceert, waaronder superoxide (O2 2), singlet zuurstof (1O2), peroxylradicaal (ROO ), stikstofmonoxide (NO ), waterstofperoxide (H2O2) en dimethylarsineperoxylradicalen [( CH3)2AsOO ].57�59 Arseen(III)-verbindingen kunnen antioxidant-enzymen remmen, met name de GSH-afhankelijke enzymen, zoals glutathion-S-transferasen (GST's), glutathionperoxidase (GSH-Px) en GSH-reductase, via binding - aan hun sulfhydryl (�SH) groepen.60,61
Lood verhoogt lipideperoxidatie. 62 Significante afname van de activiteit van weefsel SOD en hersen-GPx zijn gemeld na blootstelling aan lood. 63,64 Vervanging van zink, dat als een co-factor fungeert voor veel enzymen door lood, leidt tot inactivatie van dergelijke enzymen. Loodblootstelling kan remming van GST veroorzaken door weefselthiolen te beïnvloeden.
ROS gegenereerd door metaal-gekatalyseerde reacties kunnen DNA-basen modificeren. Drie basenubstituties, G / C, G / T en C / T, kunnen optreden als gevolg van oxidatieve schade door metaalionen, zoals Fe21, Cu21 en Ni21. Reid et al65 toonden aan dat G / C voornamelijk werd geproduceerd door Fe21 terwijl C / T-substitutie werd uitgevoerd door Cu21 en Ni21.
ANTIOXIDANTEN
Het menselijk lichaam is uitgerust met een verscheidenheid aan antioxidanten die dienen als tegengewicht voor het effect van oxidanten. Voor alle praktische doeleinden kunnen deze worden onderverdeeld in 2-categorieën: enzymatisch (tabel 2) en niet-enzymatisch (tabel 3).
Enzymatische antioxidanten
De belangrijkste enzymatische antioxidanten van de longen zijn SODs (EC 1.15.1.11), catalase (EC 1.11.1.6) en GSH-Px (EC 1.11.1.9). Naast deze belangrijke enzymen zijn ook andere antioxidanten gevonden, waaronder heem-oxygenase-1 (EC 1.14.99.3) en redox-eiwitten, zoals thioredoxines (TRXs, EC 1.8.4.10), peroxiredoxins (PRXs, EC 1.11.1.15) en glutaredoxins. spelen cruciale rollen in de pulmonale antioxidantafweer.
Omdat superoxide de primaire ROS is die uit verschillende bronnen wordt geproduceerd, is de dismutatie door SOD van primair belang voor elke cel. Alle 3-vormen van SOD, dat wil zeggen CuZn-SOD, Mn-SOD en EC-SOD, worden op grote schaal in de menselijke long tot expressie gebracht. Mn-SOD is gelokaliseerd in de matrix van de mitochondria. EC-SOD is voornamelijk gelokaliseerd in de extracellulaire matrix, vooral in gebieden die grote hoeveelheden type I collageenvezels bevatten en rond pulmonaire en systemische vaten. Het is ook gedetecteerd in het bronchiale epitheel, alveolair epitheel en alveolaire macrofagen. 66,67 Over het algemeen wordt algemeen aangenomen dat CuZn-SOD en Mn-SOD fungeren als bulkopruimers van superoxide-radicalen. Het relatief hoge EC-SOD-niveau in de long met zijn specifieke binding aan de extracellulaire matrixcomponenten kan een fundamenteel bestanddeel van de bescherming van de longmatrix vertegenwoordigen.68
H2O2 dat wordt geproduceerd door de werking van SODs of de werking van oxidasen, zoals xanthine oxidase, wordt door catalase en de GSH-Px tot water gereduceerd. Catalase bestaat als een tetra- meer samengesteld uit 4 identieke monomeren, die elk een heemgroep op de actieve plaats bevatten. Degradatie van H2O2 wordt bereikt via de omzetting tussen 2-conformaties van catalase-ferricatalase (ijzer gecoördineerd met water) en verbinding I (ijzer gecomplexeerd met een zuurstofatoom). Catalase bindt ook NADPH als een reducerend equivalent om oxidatieve inactivatie van het enzym (vorming van verbinding II) door H2O2 te voorkomen, omdat het wordt gereduceerd tot water.69
Enzymen in de redoxcyclus die verantwoordelijk zijn voor de reductie van H2O2 en lipidehydroperoxiden (gegenereerd als een resultaat van membraanperoxidatie van membranen) omvatten de GSH-Pxs.70 De GSH-Px's zijn een familie van tetramere enzymen die het unieke aminozuur selenocysteïne in de actieve locaties en gebruiken thiols met een laag moleculair gewicht, zoals GSH, om H2O2 en lipide peroxiden te reduceren tot de overeenkomstige alcoholen. Vier GSH-Px's zijn beschreven, gecodeerd door verschillende genen: GSH-Px-1 (cellulair GSH-Px) is alomtegenwoordig en vermindert H2O2 en vetzuurperoxiden, maar niet veresterde peroxyllipiden. 71 Veresterde lipiden worden verminderd door membraangebonden GSH -Px-4 (fosfolipide hydroperoxide GSH-Px), die verschillende thiolen met een laag molecuulgewicht als reducerende equivalenten kan gebruiken. GSH-Px-2 (gastro-intestinale GSH-Px) is gelokaliseerd in gastro-intestinale epitheelcellen waar het dient om peroxiden in de voeding te verminderen. 72 GSH-Px-3 (extracellulair GSH-Px) is het enige lid van de GSH-Px-familie dat zich in het extracellulaire compartiment en wordt aangenomen dat het een van de belangrijkste extracellulaire antioxidant-enzymen in zoogdieren is. Hiervan wordt extracellulair GSH-Px het meest onderzocht in de menselijke long. 73
Daarnaast is de verwijdering van H2O2 nauw verbonden met verschillende thiol-bevattende enzymen, namelijk TRXs (TRX1 en TRX2), thioredoxine-reductasen (EC 1.8.1.9) (TRR's), PRX's (die thioredoxine peroxidasen zijn) en glutaredoxins.74
Twee TRX's en TRR's zijn gekarakteriseerd in menselijke cellen, die aanwezig zijn in zowel cytosol als mitochondriën. In de long worden TRX en TRR tot expressie gebracht in bronchiaal en alveolair epitheel en macrofagen. Zes verschillende PRXs zijn gevonden in menselijke cellen, verschillend in hun ultrastructurele compartimentalisatie. Experimentele studies hebben het belang van PRX VI in de bescherming van alveolair epitheel onthuld. De menselijke long brengt alle PRXs tot expressie in bronchiaal epitheel, alveolair epitheel en macrofagen. 75 PRX V is recentelijk gebleken te functioneren als een peroxynitrietreductase, 76, wat betekent dat het kan functioneren als een potentiële beschermende verbinding in de ontwikkeling van ROS-gemedieerde longbeschadiging .77
Gemeenschappelijk voor deze antioxidanten is de eis van NADPH als een reducerend equivalent. NADPH onderhoudt katalase in de actieve vorm en wordt gebruikt als een co-factor door TRX en GSH-reductase (EC 1.6.4.2), dat GSSG omzet in GSH, een cosubstraat voor de GSH-Px's. Intracellulair NADPH wordt op zijn beurt gegenereerd door de reductie van NADP1 door glucose-6-fosfaatdehydrogenase, het eerste en snelheidsbeperkende enzym van de pen-tosefosfaatroute, tijdens de omzetting van glucose-6-fosfaat naar 6-fosfogluconolacton. Door NADPH te genereren, is glucose-6-fosfaatdehydrogenase een cruciale bepalende factor voor de cytosolische GSH-bufferende capaciteit (GSH / GSSG) en kan daarom worden beschouwd als een essentiële, regulerende antioxidante enzyme.78,79
GST's (EC 2.5.1.18), een andere familie van antioxidant-enzymen, deactiveren secundaire metabolieten, zoals onverzadigde aldehyden, epoxiden en hydroperoxiden. Er zijn drie belangrijke families van GST's beschreven: cytosolische GST, mitochondriale GST,80,81 en membraan-geassocieerde microsomale GST die een rol speelt in het eicosanoïde- en GSH-metabolisme.82 Zeven klassen van cytosolische GST worden geïdentificeerd in zoogdieren, aangeduid als Alpha, Mu, Pi, Sigma, Theta, Omega en Zeta.83 Tijdens niet-gestresste omstandigheden interageren klasse Mu en Pi GST's met kinasen Ask86 en JNK, respectievelijk, en remmen deze kinasen.1 Het is aangetoond dat GSTP87 dissocieert van JNK als reactie op oxidatieve stress.89 GSTP1 interageert ook fysiek met PRX VI en leidt tot herstel van PRX-enzymactiviteit via glutathionylering van het geoxideerde eiwit.89
Niet-enzymatische antioxidanten
Niet-enzymatische antioxidanten omvatten verbindingen met een laag moleculair gewicht, zoals vitaminen (vitamine C en E), b-caroteen, urinezuur en GSH, een tripeptide (Lg-glutamyl-L-cysteinyl-L-glycine) dat een thiol ( sulfhydryl) groep.
Vitamine C (ascorbinezuur)
Wateroplosbare vitamine C (ascorbinezuur) biedt intracellulaire en extracellulaire waterfase-antioxidantcapaciteit voornamelijk door zuurstofvrije radicalen op te ruimen. Het zet vitamine E-vrije radicalen terug naar vitamine E. Zijn plasmaspiegels bleken af te nemen met de leeftijd. 91,92
Vitamine E (a-tocoferol)
In lipiden oplosbare vitamine E is geconcentreerd in de hydrofobe inwendige plaats van het celmembraan en is de belangrijkste afweer tegen oxidant-geïnduceerde membraanbeschadiging. Vitamine E doneert elektron aan peroxylradicaal, dat wordt geproduceerd tijdens lipideperoxidatie. a-Tocoferol is de meest actieve vorm van vitamine E en de belangrijkste membraangebonden antioxidant in de cel. Vitamine E triggert apoptose van kankercellen en remt de formaties van vrije radicalen. 93
Glutathion
GSH is zeer overvloedig in alle celcompartimenten en is de belangrijkste oplosbare antioxidant. De GSH / GSSG-ratio is een belangrijke bepalende factor voor oxidatieve stress. GSH toont zijn antioxiderende effecten op verschillende manieren. 94 Het ontgift waterstofperoxide en lipidenperoxiden via de werking van GSH-Px. GSH doneert zijn elektron aan H2O2 om het te reduceren tot H2O en O2. GSSG wordt opnieuw gereduceerd tot GSH door GSH-reductase dat NAD (P) H als de elektronendonor gebruikt. GSH-Px's zijn ook belangrijk voor de bescherming van celmembraan door lipideperoxidatie. Gereduceerde glutathion doneert protonen aan membraanlipiden en beschermt ze tegen oxidantaanvallen.95
GSH is een co-factor voor verschillende ontgiftende enzymen, zoals GSH-Px en transferase. Het heeft een rol bij het omzetten van vitamine C en E naar hun actieve vormen. GSH beschermt cellen tegen apoptose door interactie met proapoptotische en anti-apoptotische signaalroutes. 94 Het reguleert en activeert ook verschillende transcriptiefactoren, zoals AP-1, NF-KB en SP-1.
Carotenoïden (b-caroteen)
Carotenoïden zijn pigmenten die in planten worden aangetroffen. In de eerste plaats is gebleken dat b-caroteen reageert met peroxyl (ROO), hydroxyl (OH) en superoxide (O22.) Radicalen. 96 Carotenoïden tonen hun antioxiderende effecten bij lage partiële zuurstofdruk maar kunnen pro-oxidanteffecten hebben bij hogere zuurstof concentration.97 Zowel carotenoïden als retinoïnezuren (RA's) zijn in staat transcriptiefactoren te reguleren. 98 b-caroteen remt de door oxidant geïnduceerde NF-KB-activering en interleukine (IL) -6 en tumor necrosefactor-een productie. Carotenoïden beïnvloeden ook apoptose van cellen. Antiproliferatieve effecten van RA zijn in verschillende onderzoeken aangetoond. Dit effect van RA wordt voornamelijk gemedieerd door retinezuurreceptoren en varieert tussen celtypen. In borstcarcinoomcellen bleek retinolzuurreceptor remming van de groei te veroorzaken door het induceren van celcyclusstilstand, apoptose of both.99,100
HET EFFECT VAN OXIDATIEVE STRESS: GENETISCHE, FYSIOLOGISCHE EN BIOCHEMISCHE MECHANISMEN
Oxidatieve stress treedt op wanneer de balans tussen antioxidanten en ROS wordt verstoord door hetzij uitputting van antioxidanten of accumulatie van ROS. Wanneer oxidatieve stress optreedt, proberen cellen de oxiderende effecten tegen te gaan en de redoxbalans te herstellen door activering of silencing van genen die coderen voor defensieve enzymen, transcriptiefactoren en structurele eiwitten. 101,102 Verhouding tussen geoxideerd en gereduceerd glutathion (2GSH / GSSG) is één van de belangrijke determinanten van oxidatieve stress in het lichaam. Een hogere productie van ROS in het lichaam kan de DNA-structuur veranderen, resulteren in modificatie van eiwitten en lipiden, activering van verschillende stress-geïnduceerde transcriptiefactoren en de productie van pro-inflammatoire en anti-inflammatoire cytokines.
Effecten van oxidatieve stress op DNA
ROS kan op verschillende manieren leiden tot DNA-modificaties, wat afbraak van basen, enkel- of dubbelstrengs DNA-breuken, purine, pyrimidine of aan suiker gebonden modificaties, mutaties, deleties of translocaties en verknoping met eiwitten omvat. De meeste van deze DNA-modificaties (Fig. 1) zijn zeer relevant voor carcinogenese, veroudering en neurodegeneratieve, cardiovasculaire en auto-immuunziekten. Tabaksrook, redox-metalen en niet-redox-metalen, zoals ijzer, cadmium, chroom en arseen, zijn ook betrokken bij carcinogenese en veroudering door het genereren van vrije radicalen of het binden aan thiolgroepen. Vorming van 8-OH-G is de bekendste DNA-schade die optreedt via oxidatieve stress en is een potentiële biomarker voor carcinogenese.
Promotorregio's van genen bevatten consensussequenties voor transcriptiefactoren. Deze bindingsplaatsen voor transcriptiefactoren bevatten GC-rijke sequenties die vatbaar zijn voor oxidantaanvallen. Vorming van 8-OH-G-DNA in bindingsplaatsen voor transcriptiefactoren kan de binding van transcriptiefactoren wijzigen en zo de expressie van verwante genen veranderen, zoals is aangetoond voor AP-1- en Sp-1-doelsequenties.103 Naast 8-OH-G, Van 8,59 -cyclo-29-deoxyadenosine (cyclo-dA) is ook aangetoond dat het de transcriptie van een reportergen in een celsysteem remt als het zich in een TATA-box bevindt.104 Het TATA-bindende eiwit initieert transcriptie door de buiging van DNA te veranderen . De binding van TATA-bindend eiwit kan worden aangetast door de aanwezigheid van cyclo-dA.
Oxidatieve stress veroorzaakt instabiliteit van regio's met microsatelliet (korte tandemherhalingen). Redox-actieve metaalionen, hydroxylradicalen verhogen de instabiliteit van microsatelliet. 105 Hoewel enkelstrengige DNA-breuken veroorzaakt door oxidantschade gemakkelijk door cellen kunnen worden getolereerd, kunnen dubbelstrengige DNA-breuken geïnduceerd door ioniserende straling een significante bedreiging vormen voor de celoverleving.106
Methylatie op CpG-eilanden in DNA is een belangrijk epigenetisch mechanisme dat kan leiden tot gene silencing. Oxidatie van 5-MeCyt tot 5-hydroxymethyluracil (5-OHMeUra) kan plaatsvinden via deaminerings- / oxidatiereacties van thymine of 5-hydroxymethylcytosine-tussenproducten. 107 Naast de modulerende genexpressie lijkt DNA-methylatie ook de chromatine-organisatie te beïnvloeden.108 Afwijkende DNA-methylatiepatronen geïnduceerd door oxidatieve aanvallen beïnvloeden ook de DNA-herstelactiviteit.
Effecten van oxidatieve stress op lipiden
ROS kan lipideperoxidatie induceren en de opstelling van de membraanlipidendubbellaag verstoren, wat membraangebonden receptoren en enzymen kan inactiveren en de weefselpermeabiliteit kan verhogen.109 Producten van lipideperoxidatie, zoals MDA en onverzadigde aldehyden, zijn in staat veel cellulaire eiwitten te inactiveren door eiwitkruising te vormen. -koppelingen.110�112 4-Hydroxy-2-nonenal veroorzaakt uitputting van intracellulair GSH en induceert peroxideproductie,113,114 activeert epidermale groeifactorreceptor,115 en induceert fibronectineproductie.116 Lipideperoxidatieproducten, zoals isoprostanen en thiobarbituurzuurreactieve stoffen , zijn gebruikt als indirecte biomarkers van oxidatieve stress, en verhoogde niveaus werden aangetoond in het uitgeademde ademcondensaat of bronchoalveolaire lavagevloeistof of long van patiënten met chronische obstructieve longziekte of rokers.117�119
Effecten van oxidatieve stress op proteïnen
ROS kan fragmentatie van de peptideketen, wijziging van de elektrische lading van eiwitten, verknoping van eiwitten en oxidatie van specifieke aminozuren veroorzaken en daardoor leiden tot verhoogde gevoeligheid voor proteolyse door afbraak door specifieke proteasen.120 Cysteïne- en methionineresiduen in eiwitten zijn bijzonder gevoeliger voor oxidatie.121 Oxidatie van sulfhydrylgroepen of methionineresiduen van eiwitten veroorzaken conformationele veranderingen, eiwitontvouwing en afbraak.8,121�123 Enzymen met metalen op of dichtbij hun actieve plaatsen zijn bijzonder gevoeliger voor door metaal gekatalyseerde oxidatie. Het is aangetoond dat oxidatieve modificatie van enzymen hun activiteiten remt.124,125
In sommige gevallen kan specifieke oxidatie van eiwitten plaatsvinden. Methionine kan bijvoorbeeld geoxideerd methioninesulfoxide 126 en fenylalanine tot o-tyrosine127 zijn; sulfhydrylgroepen kunnen worden geoxideerd om disulfidebindingen te vormen; 128 en carbonylgroepen kunnen in de zijketens van eiwitten worden geïntroduceerd. Gamma-stralen, metaalgekatalyseerde oxidatie, HOCl en ozon kunnen de vorming van carbonylgroepen veroorzaken.129
Effecten van oxidatieve stress op signaaltransductie
ROS kan expressie induceren van verschillende genen die betrokken zijn bij signaaltransductie.1,130 Een hoge verhouding voor GSH/GSSG is belangrijk voor de bescherming van de cel tegen oxidatieve schade. Verstoring van deze verhouding veroorzaakt activering van redox-gevoelige transcriptiefactoren, zoals NF-kB, AP-1, nucleaire factor van geactiveerde T-cellen en hypoxie-induceerbare factor 1, die betrokken zijn bij de ontstekingsreactie. Activering van transcriptiefactoren via ROS wordt bereikt door signaaltransductiecascades die de informatie van buiten naar binnen in de cel overbrengen. Tyrosinekinasereceptoren, de meeste groeifactorreceptoren, zoals epidermale groeifactorreceptor, vasculaire endotheliale groeifactorreceptor en receptor voor van bloedplaatjes afgeleide groeifactor, eiwittyrosinefosfatasen en serine/threoninekinasen zijn doelwitten van ROS.131�133 Extracellulaire signaalgereguleerde kinasen, JNK en p38, die de leden zijn van de mitogeen-geactiveerde eiwitkinasefamilie en betrokken zijn bij verschillende celprocessen, waaronder proliferatie, differentiatie en apoptose, kunnen ook worden gereguleerd door oxidanten.
Onder omstandigheden van oxidatieve stress ondergaan cysteïneresiduen in de DNA-bindingsplaats van c-Jun, sommige AP-1-subeenheden en remmende kB-kinase reversibele S-glutathiolering. Van glutaredoxine en TRX is gemeld dat ze een belangrijke rol spelen bij de regulatie van redoxgevoelige signaalroutes, zoals NF-kB en AP-1, p38 mitogeen-geactiveerd proteïnekinase en JNK.134�137
NF-kB kan worden geactiveerd als reactie op oxidatieve stressomstandigheden, zoals ROS, vrije radicalen en UV-straling.138 Fosforylering van IkB maakt NF-kB vrij en stelt het in staat de kern binnen te gaan om gentranscriptie te activeren.139 Een aantal kinasen heeft er is gemeld dat ze IkB's fosforyleren op de serineresiduen. Deze kinasen zijn het doelwit van oxidatieve signalen voor activatie van NF-kB.140 Reductiemiddelen versterken de DNA-binding van NF-kB, terwijl oxidatiemiddelen de DNA-binding van NF-kB remmen. TRX kan 2 tegengestelde acties uitoefenen bij de regulatie van NF-kB: in het cytoplasma blokkeert het de afbraak van IkB en remt het de activering van NF-kB, maar verbetert het de binding van NF-kB-DNA in de kern.141 Activering van NF-kB via oxidatiegerelateerde afbraak van IkB resulteert in de activering van verschillende genen die verband houden met de verdediging van antioxidanten. NF-kB reguleert de expressie van verschillende genen die deelnemen aan de immuunrespons, zoals IL-1b, IL-6, tumornecrosefactor-a, IL-8 en verschillende adhesiemoleculen.142,143 NF-kB reguleert ook angiogenese en proliferatie en differentiatie van cellen.
AP-1 wordt ook gereguleerd door redox-status. In aanwezigheid van H2O2 kunnen sommige metaalionen de AP-1 activeren. Toename in de verhouding van GSH / GSSG verhoogt AP-1-binding, terwijl GSSG de DNA-binding van AP-1.144 remt DNA-binding van het Fos / Jun-heterodimeer wordt verhoogd door de reductie van één geconserveerd cysteïne in het DNA-bindingsdomein van elk van de eiwitten, 145, terwijl DNA-binding van AP-1 kan worden geremd door GSSG in veel celtypen, wat suggereert dat vorming van disulfidebindingen door cysteïne-residuen AP-1-DNA-binding remt. 146,147 Signaaltransductie via oxidatieve stress is samengevat in Figuur 2.
CONCLUSIES
Oxidatieve stress kan het gevolg zijn van overproductie van ROS door metabolische reacties die zuurstof gebruiken en de balans ertussen verschuiven oxidant /anti-oxidant statussen ten gunste van de oxidatiemiddelen. ROS worden geproduceerd door cellulaire metabolische activiteiten en omgevingsfactoren, zoals luchtverontreinigende stoffen of sigarettenrook. ROS zijn zeer reactieve moleculen vanwege ongepaarde elektronen in hun structuur en reageren met verschillende biologische macromoleculen in cellen, zoals koolhydraten, nucleïnezuren, lipiden en eiwitten, en veranderen hun functies. ROS beïnvloedt ook de expressie van verschillende genen door opregulering van redox-gevoelige transcriptiefactoren en chromatine remodellering via wijziging in histonacetylering / deacetylatie. Regulering van de redox-toestand is van cruciaal belang voor cellevensvatbaarheid, activering, proliferatie en orgaanfunctie.
REFERENTIES
1. Valko M, Rhodes CJ, Moncol J, Izakovic M, Mazur M. Vrije radicalen, metalen en antioxidanten bij door oxidatieve stress veroorzaakte kanker. Chem Biol-interactie. 2006;160:1�40.
2. Halliwell B, Gutteridge JMC. Vrije radicalen in biologie en geneeskunde. 3rd ed. New York: Oxford University Press; 1999.
3. Marnett LJ. Lipideperoxidatie dDNA-schade door malondialdehyde. Mutat Res. 1999;424:83�95.
4. Siems WG, Grune T, Esterbauer H. 4-Hydroxynonenal-vorming tijdens ischemie en reperfusie van de dunne darm van de rat. �Life Science. 1995;57:785�789.
5. Stadtman ER. De rol van oxidantsoorten bij veroudering. Curr Med Chem. 2004;11:1105�1112.
6. Wang MIJN, Dhingra K, Hittelman WN, Liehr JG, deAndrade M, Li DH. Door lipideperoxidatie geïnduceerde vermoedelijke malondialdehyde-DNA-adducten in menselijk borstweefsel. Kanker Epidemiol Biomarkers Vorige. 1996;5:705�710.
7. Jenner P. Oxidatieve stress bij de ziekte van Parkinson. Ann Neurol. 2003;53: S26�S36.
8. Lyras L, Cairns NJ, Jenner A, Jenner P, Halliwell B. Een beoordeling van oxidatieve schade aan eiwitten, lipiden en DNA in de hersenen van patiënten met de ziekte van Alzheimer. J Neurochem. 1997;68:2061�2069.
9. Sayre LM, Smith MA, Perry G. Chemie en biochemie van oxidatieve stress bij neurodegeneratieve ziekten. Curr Med Chem. 2001;8:721�738.
10. Toshniwal PK, Zarling EJ. Bewijs voor verhoogde lipideperoxidatie bij multiple sclerose. Neurochem Res. 1992;17:205�207.
11. Dhalla NS, Temsah RM, Netticadan T. De rol van oxidatieve stress bij hart- en vaatziekten. J Hypertens. 2000;18:655�673.
12. Kasparova S, Brezova V, Valko M, Horecky J, Mlynarik V, et al. Studie van de oxidatieve stress in een rattenmodel van chronische hersenhyperfusie. Neurochem Int. 2005;46:601�611.
13. Kerr S, Brosnan MJ, McIntyre M, Reid JL, Dominiczak AF, Hamilton CA. De productie van superoxide-anionen is verhoogd in een model van genetische hypertensie: rol van het endotheel. Hypertensie. 1999;33:1353�1358.
14. Kukreja RC, Hess ML. Het zuurstof-vrije-radicalensysteem: van vergelijkingen via membraan-eiwitinteracties tot cardiovasculair letsel en bescherming. Cardiovasc Res. 1992;26:641�655.
15. Asami S, Manabe H, Miyake J, Tsurudome Y, Hirano T, et al. Het roken van sigaretten veroorzaakt een toename van oxidatieve DNA-schade, 8-hydroxydeoxyguanosine, op een centrale plek in de menselijke long. Kankerverwekkendheid. 1997;18:1763-1766.
16. Andreadis AA, Hazen SL, Comhair SA, Erzurum SC. Oxidatieve en nitrosatieve gebeurtenissen bij astma. Gratis radicaal Biol Med. 2003;35:213�225.
17. Comhair SA, Ricci KS, Arroliga M, Lara AR, Dweik RA, et al. Correlatie van systemische superoxide-dismutasedeficiëntie met luchtwegobstructie bij astma. Am J Respir Crit Care Med. 2005;172:306�313.
18. Comhair SA, Xu W, Ghosh S, Thunnissen FB, Almasan A, et al. Superoxide-dismutase-inactivatie in pathofysiologie van remodellering en reactiviteit van astmatische luchtwegen. Ben J Pathol. 2005;166:663�674.
19. Dut R, Dizdar EA, Birben E, Sackesen C, Soyer OU, Besler T, Kalayci O. Oxidatieve stress en de determinanten ervan in de luchtwegen van kinderen met astma. Allergie. 2008;63:1605�1609.
20. Ercan H, Birben E, Dizdar EA, Keskin O, Karaaslan C, et al. Oxidatieve stress en genetische en epidemiologische determinanten van oxidatieve schade bij astma bij kinderen. J Allergie Clin Immunol. 2006;118:1097�1104.
21. Fitzpatrick AM, Teague WG, Holguin F, Yeh M, Brown LA. Onderzoeksprogramma voor ernstig astma. De homeostase van glutathion in de luchtwegen is veranderd bij kinderen met ernstige astma: bewijs voor oxidatieve stress. J Allergie Clin Immunol. 2009;123:146�152.
22. Miller DM, Buettner GR, Aust SD. Overgangsmetalen als katalysatoren van "auto-oxidatie"-reacties. Gratis radicaal Biol Med. 1990;8:95�108.
23. Dupuy C, Virion A, Ohayon R, Kaniewski J, D�me D, Pommier J. Mechanisme van vorming van waterstofperoxide gekatalyseerd door NADPH-oxidase in plasmamembraan van de schildklier. J Biol Chem. 1991;266:3739�3743.
24. Griffel DN. De rol van xanthine-oxidase en granulocyten bij ischemie-reperfusieschade. Ben J Physiol. 1988;255:H1269�H1275.
25. Fenton HJH. Oxidatie van wijnsteenzuur in aanwezigheid van ijzer. JChem Soc. 1984;65:899�910.
26. Haber F, Weiss JJ. De katalytische ontleding van waterstofperoxide door ijzerzouten. Proc R Soc Lond Ser A. 1934;147:332�351.
27. Liochev SI, Fridovich I. De Haber�Weiss fietste 70 jaar later: een alternatieve kijk. Redox Rep. 2002;7:55�57.
28. Klebanoff SJ. Myeloperoxidase: vriend en vijand. J Leukoc Biol. 2005;77:598�625.
29. Whiteman M, Jenner A, Halliwell B. Hypochloorzuur-geïnduceerde basemodificaties in geïsoleerd kalfsthymus-DNA. Chem Res Toxicol. 1997;10:1240�1246.
30. Kulcharyk PA, Heinecke JW. Hypochloorzuur geproduceerd door het myeloperoxidasesysteem van menselijke fagocyten induceert covalente verknopingen tussen DNA en eiwit. Biochemie. 2001;40:3648�3656.
31. Brennan ML, Wu W, Fu X, Shen Z, Song W, et al. Een verhaal over twee controverses: het definiëren van zowel de rol van peroxidasen bij de vorming van nitrotyrosine in vivo met behulp van eosinofiele peroxidase en myeloperoxidasedeficiënte muizen, als de aard van door peroxidase gegenereerde reactieve stikstofsoorten. J Biol Chem. 2002;277:17415�17427.
32. Denzler KL, Borchers MT, Crosby JR, Cieslewicz G, Hines EM, et al. Uitgebreide degranulatie van eosinofielen en peroxidase-gemedieerde oxidatie van luchtwegeiwitten komen niet voor in een ovalbumine-uitdagingsmodel van longontsteking bij muizen. J Immunol. 2001;167:1672�1682.
33. van Dalen CJ, Winterbourn CC, Senthilmohan R, Kettle AJ. Nitriet als substraat en remmer van myeloperoxidase. Implicaties voor nitratie en hypochloorzuurproductie op ontstekingsplaatsen. J Biol Chem. 2000;275:11638�11644.
34. Wood LG, Fitzgerald DA, Gibson PG, Cooper DM, Garg ML. Lipidenperoxidatie zoals bepaald door plasma-isoprostanen is gerelateerd aan de ernst van de ziekte bij milde astma. Lipiden. 2000;35:967�974.
35. Montuschi P, Corradi M, Ciabattoni G, Nightingale J, Kharitonov SA, Barnes PJ. Verhoogd 8-isoprostaan, een marker van oxidatieve stress, in uitgeademd condensaat van astmapatiënten. Am J Respir Crit Care Med. 1999;160:216�220.
36. Kerk DF, Pryor WA. Vrije radicalenchemie van sigarettenrook en de toxicologische implicaties ervan. Milieugezondheidsperspectief. 1985;64:111�126.
37. Hiltermann JT, Lapperre TS, van Bree L, Steerenberg PA, Brahim JJ, et al. Ozon-geïnduceerde ontsteking beoordeeld in sputum en bronchiale lavagevloeistof van astmapatiënten: een nieuw niet-invasief hulpmiddel in epidemiologische studies over luchtverontreiniging en astma. Gratis radicaal Biol Med. 1999;27:1448�1454.
38. Nachtegaal JA, Rogers DF, Barnes PJ. Effect van ingeademde ozon op uitgeademd stikstofmonoxide, longfunctie en geïnduceerd sputum bij normale en astmatische personen. Thorax. 1999;54:1061�1069.
39. Cho AK, Sioutas C, Miguel AH, Kumagai Y, Schmitz DA, et al. Redox-activiteit van fijnstof in de lucht op verschillende locaties in het Los Angeles Basin. Omgeving Res. 2005;99:40�47.
40. Comhair SA, Thomassen MJ, Erzurum SC. Differentiële inductie van extracellulair glutathionperoxidase en stikstofmonoxidesynthase 2 in de luchtwegen van gezonde personen die zijn blootgesteld aan 100% O (2) of sigarettenrook. Am J Adem Cell Mol Biol. 2000;23:350�354.
41. Matthay MA, Geiser T, Matalon S, Ischiropoulos H. Oxidant-gemedieerde longbeschadiging bij het acute respiratory distress syndrome. Crit Care Med. 1999;27:2028�2030.
42. Biaglow JE, Mitchell JB, Held K. Het belang van peroxide en superoxide in de röntgenreactie. Int J Radiat Oncol Biol Phys. 1992;22:665�669.
43. Chiu SM, Xue LY, Friedman LR, Oleinick NL. Door koperionen gemedieerde sensibilisatie van bevestigingsplaatsen van nucleaire matrix voor ioniserende straling. Biochemie. 1993;32:6214�6219.
44. Narayanan PK, Goodwin EH, Lehnert BE. Alfadeeltjes initiëren de biologische productie van superoxide-anionen en waterstofperoxide in menselijke cellen. kanker res. 1997;57:3963�3971.
45. Tuttle SW, Varnes ME, Mitchell JB, Biaglow JE. Gevoeligheid voor chemische oxidanten en straling in CHO-cellijnen met een tekort aan oxidatieve pentosecyclusactiviteit. Int J Radiat Oncol Biol Phys. 1992;22: 671�675.
46. Guo G, Yan-Sanders Y, Lyn-Cook BD, Wang T, Tamae D, et al. Mangaan
superoxide dismutase-gemedieerde genexpressie in geïnduceerde straling
adaptieve reacties. Mol Cel Biol. 2003;23:2362�2378.
47. Azzam EI, de Toledo SM, Spitz DR, Little JB. Oxidatief metabolisme
moduleert signaaltransductie en micronucleusvorming bij omstanders
cellen van a-deeltjes bestraalde normale menselijke fibroblasten. Cancer Res.
2002; 62: 5436.
48. Leach JK, Van Tuyle G, Lin PS, Schmidt-Ullrich R, Mikkelsen RB.
Ioniserende straling-geïnduceerde, mitochondria-afhankelijke generatie van reactief
zuurstof/stikstof. kanker res. 2001;61:3894�3901.
49. Dent P, Yacoub A, Fisher PB, Hagan MP, Grant S. MAPK-routes in
straling reacties. Oncogeen. 2003;22:5885�5896.
50. Wei SJ, Botero A, Hirota K, Bradbury CM, Markovina S, et al. thioredoxine
nucleaire translocatie en interactie met redoxfactor-1 activeert de AP-1-transcriptiefactor als reactie op ioniserende straling. kanker res. 2000;60:6688�6695.
51. Cadet J, Douki T, Gasparutto D, Ravanat JL. Oxidatieve schade aan DNA: vorming, meting en biochemische kenmerken. Mutat Res. 2003;531:5�23.
52. Yokoya A, Cunniffe SM, O�Neill P. Effect van hydratatie op de inductie van strengbreuken en basislaesies in plasmide-DNA-films door gammastraling. J Am Chem Soc. 2002;124:8859�8866.
53. Janssen YM, Van Houten B, Borm PJ, Mossman BT. Cel- en weefselreacties op oxidatieve schade. Lab Invest. 1993;69:261�274.
54. Iwanaga M, Mori K, Iida T, Urata Y, Matsuo T, et al. Nucleaire factor kappa B-afhankelijke inductie van gamma-glutamylcysteïnesynthetase door ioniserende straling in menselijke T98G-glioblastoomcellen. Gratis radicaal Biol Med. 1998;24:1256�1268.
55. Stohs SJ, Bagchi D. Oxidatieve mechanismen in de toxiciteit van metaalionen. Gratis radicaal Biol Med. 1995;18:321�336.
56. Leonard SS, Harris GK, Shi X. Metaalgeïnduceerde oxidatieve stress en signaaltransductie. Gratis radicaal Biol Med. 2004;37:1921-1942.
57. Shi H, Shi X, Liu KJ. Oxidatief mechanisme van arseentoxiciteit en carcinogenese. Mol Cell Biochem. 2004;255:67�78.
58. Pi J, Horiguchi S, Sun Y, Nikaido M, Shimojo N, Hayashi T. Een mogelijk mechanisme voor de verslechtering van de vorming van stikstofmonoxide veroorzaakt door langdurige orale blootstelling aan arsenaat bij konijnen. Vrije radicaal Biol Med.2003;35:102�113.
59. Rin K, Kawaguchi K, Yamanaka K, Tezuka M, Oku N, Okada S. DNA-strengbreuken geïnduceerd door dimethylarsinezuur, een metaboliet van anorganisch arseen, worden sterk versterkt door superoxide-anionradicalen. Biol Pharm Stier. 1995;18:45�58.
60. Waalkes MP, Liu J, Ward JM, Diwan LA. Mechanismen die ten grondslag liggen aan arseencarcinogenese: overgevoeligheid van muizen die tijdens de dracht worden blootgesteld aan anorganisch arseen. Toxicologie. 2004;198:31�38.
61. Schiller CM, Fowler BA, Woods JS. Effecten van arseen op activering van pyruvaatdehydrogenase. Milieugezondheidsperspectief. 1977;19:205�207.
62. Monterio HP, Bechara EJH, Abdalla DSP. Betrokkenheid van vrije radicalen bij neurologische porfyrieën en loodvergiftiging. Mol Cell Biochem. 1991;103:73�83.
63. Tripathi RM, Raghunath R, Mahapatra S. Bloedlood en het effect ervan op de Cd-, Cu-, Zn-, Fe- en hemoglobinespiegels van kinderen. Sci Totale Omgeving. 2001;277:161�168.
64. Nehru B, Dua R. Het effect van selenium in de voeding op loodneurotoxiciteit. J Environ Pathol Toxicol Oncol. 1997;16:47�50.
65. Reid TM, Feig DI, Loeb LA. Mutagenese door metaalgeïnduceerde zuurstofradicalen. Milieugezondheidsperspectief. 1994;102(suppl. 3):57�61.
66. Kinnula VL, Crapo JD. Superoxide-dismutasen in de longen en longziekten bij de mens. Am J Respir Crit Care Med. 2003;167:1600�1619.
67. Kinnula VL. Productie en afbraak van zuurstofmetabolieten tijdens ontstekingstoestanden in de menselijke longen. Curr-geneesmiddel richt zich op ontstekingsallergie. 2005;4:465�470.
68. Zelko IN, Mariani TJ, Folz RJ. Superoxide dismutase multigene familie: een vergelijking van de CuZn-SOD (SOD1), Mn-SOD (SOD2) en EC-SOD (SOD3) genstructuren, evolutie en expressie. Gratis radicaal Biol Med. 2002;33:337�349.
69. Kirkman HN, Rolfo M, Ferraris AM, Gaetani GF. Beschermingsmechanismen van catalase door NADPH. Kinetiek en stoichiometrie. J Biol Chem. 1999;274:13908�13914.
70. Floh� L. Glutathionperoxidase. Basis levenswetenschap. 1988;49:663�668.
71. Arthur JR. De glutathionperoxidasen. Cel Mol Leven Sci. 2000;57:1825-1835.
72. Chu FF, Doroshow JH, Esworthy RS. Expressie, karakterisering en weefseldistributie van een nieuwe cellulaire seleniumafhankelijke glutathionperoxidase, GSHPx-GI. J Biol Chem. 1993;268:2571�2576.
73. Comhair SA, Bhathena PR, Farver C, Thunnissen FB, Erzurum SC. Extracellulaire glutathionperoxidase-inductie in astmatische longen: bewijs voor redox-regulatie van expressie in menselijke luchtwegepitheelcellen. FASEB J. 2001;15:70�78.
74. Gromer S, Urig S, Becker K. Het thioredoxinesysteem van wetenschap tot kliniek. Med Res Rev. 2004;24:40�89.
75. Kinnula VL, Lehtonen S, Kaarteenaho-Wiik R, Lakari E, P��kk� P, et al. Celspecifieke expressie van peroxiredoxinen in long- en longsarcoïdose bij de mens. Thorax. 2002;57:157�164.
76. Dubuisson M, Vander Stricht D, Clippe A, Etienne F, Nauser T, et al. Humaan peroxiredoxine 5 is een peroxynitrietreductase. FEBS Lett. 2004;571:161�165.
77. Holmgren A. Antioxiderende functie van thioredoxine- en glutaredoxinesystemen. Antioxide Redox-signaal. 2000;2:811�820.
78. Dickinson DA, Forman HJ. Glutathion in verdediging en signalering: lessen uit een kleine thiol. Ann NY Acad Sci. 2002;973:488�504.
79. Sies H. Glutathione en zijn rol in cellulaire functies. Gratis radicaal Biol Med. 1999;27:916�921.
80. Ladner JE, Parsons JF, Rife CL, Gilliland GL, Armstrong RN. Parallelle evolutionaire routes voor glutathiontransferasen: structuur en mechanisme van het mitochondriale klasse kappa-enzym rGSTK1-1. Biochemie. 2004;43:52�61.
81. Robinson A, Huttley GA, Booth HS, Board PG. Modellerings- en bio-informatica-onderzoeken van de humane kappa-klasse glutathiontransferase voorspellen een nieuwe derde transferase-familie met homologie met prokaryote 2-hydroxychromeen-2-carboxylaat-isomerasen. Biochem J. 2004;379:541-552.
82. Jakobsson PJ, Morgenstern R, Mancini J, Ford-Hutchinson A, Persson B. Gemeenschappelijke structurele kenmerken van MAPEGda wijdverbreide superfamilie van membraan-geassocieerde eiwitten met sterk uiteenlopende functies in het metabolisme van eicosanoïden en glutathion. Eiwit Wetenschap. 1999;8:689�692.
83. Hayes JD, DJ Pulford. De glutathione S-transferase supergenenfamilie: regulatie van GST en de bijdrage van de iso-enzymen aan kankerchemoprotectie en resistentie tegen geneesmiddelen. Crit Rev Biochem Mol Biol. 1995;30:445�600.
84. Armstrong RN. Structuur, katalytisch mechanisme en evolutie van de glutathiontransferasen. Chem Res Toxicol. 1997;10:2�18.
85. Hayes JD, McLellan LI. Glutathion en glutathion-afhankelijke enzymen vertegenwoordigen een gecoördineerd gereguleerde afweer tegen oxidatieve stress. Gratis radicale res. 1999;31:273.
86. Sheehan D, Meade G, Foley VM, Dowd CA. Structuur, functie en evolutie van glutathiontransferasen: implicaties voor classificatie van niet-zoogdierlijke leden van een oude enzymsuperfamilie. Biochem J. 2001;360:1�16.
87. Cho SG, Lee YH, Park HS, Ryoo K, Kang KW, et al. Glutathion S-transferase Mu moduleert de door stress geactiveerde signalen door apoptose-signaalregulerende kinase 1 te onderdrukken. J Biol Chem. 2001;276:12749�12755.
88. Dorion S, Lambert H, Landry J. Activering van de p38-signaleringsroute door hitteschok omvat de dissociatie van glutathion S-transferase Mu van Ask1. J Biol Chem. 2002;277:30792�30797.
89. Adler V, Yin Z, Fuchs SY, Benezra M, Rosario L, et al. Regulering van JNK-signalering door GSTp. EMBO J. 1999;18:1321�1334.
90. Manevich Y, Feinstein SI, Fisher AB. Activering van het antioxidant-enzym 1-CYS peroxiredoxine vereist glutathionylering gemedieerd door heterodimerisatie met pGST. Proc Natl Acad Sci US A. 2004;101:3780�3785.
91. Bunker VW. Vrije radicalen, antioxidanten en veroudering. Med Lab Science. 1992;49:299�312.
92. Mezzetti A, Lapenna D, Romano F, Costantini F, Pierdomenico SD, et al. Systemische oxidatieve stress en de relatie met leeftijd en ziekte. J Am Geriatr Soc. 1996;44:823�827.
93. White E, Shannon JS, Patterson RE. Verband tussen vitamine en
gebruik van calciumsupplementen en darmkanker. Kanker Epidemiol Biomarkers Vorige. 1997;6:769�774.
94. Masella R, Di Benedetto R, Vari R, Filesi C, Giovannini C. Nieuwe mechanismen van natuurlijke antioxidantverbindingen in biologische systemen: betrokkenheid van glutathion en glutathion-gerelateerde enzymen. J Nutr Biochem. 2005;16:577�586.
95. Curello S, Ceconi C, Bigoli C, Ferrari R, Albertini A, Guarnieri C. Veranderingen in de cardiale glutathionstatus na ischemie en reperfusie. ervaring. 1985;41:42�43.
96. El-Agamey A, Lowe GM, McGarvey DJ, Mortensen A, Phillip DM, Truscott TG. Carotenoïde radicaalchemie en antioxiderende/pro-oxidante eigenschappen. Arch Biochem Biophys. 2004;430:37�48.
97. Rice-Evans CA, Sampson J, Bramley PM, Holloway DE. Waarom verwachten we dat carotenoïden in vivo antioxidanten zijn? Gratis radicale res. 1997;26:381�398.
98. Niles RM. Signaalroutes bij retinoïde chemopreventie en behandeling van kanker. Mutat Res. 2004;555:81�96.
99. Donato LJ, Noy N. Onderdrukking van borstcarcinoomgroei door retinoïnezuur: proapoptotische genen zijn doelen voor retinoïnezuurreceptor en cellulaire retinoïnezuurbindende proteïne II-signalering. kanker res. 2005;65:8193�8199.
100. Niizuma H, Nakamura Y, Ozaki T, Nakanishi H, Ohira M, et al. Bcl-2 is een belangrijke regulator voor de door retinoïnezuur geïnduceerde apoptotische celdood bij neuroblastoom. Oncogeen. 2006;25:5046�5055.
101. Dalton TP, Shertzer HG, Puga A. Regulatie van genexpressie door reactieve zuurstof. Ann Rev Pharmacol Toxicol. 1999;39:67�101.
102. Scandalios JG. Genomische reacties op oxidatieve stress. In: Meyers RA, uitg. Encyclopedie van moleculaire celbiologie en moleculaire geneeskunde. Deel 5. 2e druk. Weinheim, Duitsland: Wiley-VCH; 2004: 489�512.
103. Ghosh R, Mitchell DL. Effect van oxidatieve DNA-schade in promotorelementen op transcriptiefactorbinding. Nucleïnezuren Res. 1999;27:3213�3218.
104. Marietta C, Gulam H, Brooks PJ. Een enkele 8-cyclo-50-deoxyadenosine-laesie in een TATA-box voorkomt binding van het TATA-bindende eiwit en vermindert de transcriptie in vivo sterk. DNA-reparatie (Amst). 20;2002:1�967.
105. Jackson AL, Chen R, Loeb LA. Inductie van instabiliteit van microsatelliet
door oxidatieve DNA-schade. Proc Natl Acad Sci VS A. 1998;95:12468�12473.
106. Caldecott KW. Eiwit-eiwitinteracties tijdens het herstel van enkelstrengs DNA-breuken bij zoogdieren. Biochem Soc Trans. 2003;31:247�251.
107. Cooke MS, Evans MD, Dizdaroglu M, Lunec J. Oxidatieve DNA-schade: mechanismen, mutatie en ziekte. FASEB J. 2003;17:1195�1214.
108. Jones PL, Wolffe AP. Relaties tussen chromatine-organisatie en DNA-methylatie bij het bepalen van genexpressie. Semin Kanker Biol. 1999;9:339�347.
109. Girotti AW. Mechanismen van lipideperoxidatie. J Vrije Radicalen Biol Med. 1985;1:87�95.
110. Siu GM, Draper HH. Metabolisme van malonaldehyde in vivo en in vitro. Lipiden. 1982;17:349�355.
111. Esterbauer H, Koller E, Slee RG, Koster JF. Mogelijke betrokkenheid van het lipideperoxidatieproduct 4-hydroxynonenal bij de vorming van fluorescerende chromolipiden. Biochem J. 1986;239:405-409.
112. Hagihara M, Nishigaki I, Maseki M, Yagi K. Leeftijdsafhankelijke veranderingen in lipideperoxideniveaus in de lipoproteïnefracties van menselijk serum. J Gerontol. 1984;39:269�272.
113. Keller JN, Mark RJ, Bruce AJ, Blanc E, Rothstein JD, et al. 4- Hydroxynonenal, een aldehydeproduct van membraanlipideperoxidatie, schaadt glutamaattransport en mitochondriale functie in synaptosomen. Neurowetenschap. 1997;806:85�96.
114. Uchida K, Shiraishi M, Naito Y, Torii Y, Nakamura Y, Osawa T. Activering van stresssignaleringsroutes door het eindproduct van lipideperoxidatie. 4-hydroxy-2-nonenal is een potentiële inductor van intracellulaire peroxideproductie. J Biol Chem. 1999;274:2234�2242.
115. Suc I, Meilhac O, Lajoie-Mazenc I, Vandaele J, Jurgens G, Salvayre R, Negre-Salvayre A. Activering van EGF-receptor door geoxideerd LDL. FASEB J. 1998;12:665�671.
116. Tsukagoshi H, Kawata T, Shimizu Y, Ishizuka T, Dobashi K, Mori M. 4-Hydroxy-2-nonenal verbetert de productie van fibronectine door IMR-90 menselijke longfibroblasten, gedeeltelijk via activering van epidermale groeifactorreceptor-gekoppeld extracellulair signaal- gereguleerde kinase p44/42-route. Toxicol Appl Pharmacol. 2002;184:127�135.
117. Montuschi P, Collins JV, Ciabattoni G, Lazzeri N, Corradi M, Kharitonov SA, Barnes PJ. Uitgeademd 8-isoprostaan als een in vivo biomarker van oxidatieve stress in de longen bij patiënten met COPD en gezonde rokers. Am J Respir Crit Care Med. 2000;162:1175�1177.
118. Morrison D, Rahman I, Lannan S, MacNee W. Epitheliale permeabiliteit, ontsteking en oxidatieve stress in de luchtruimten van rokers. Am J Respir Crit Care Med. 1999;159:473�479.
119. Nowak D, Kasielski M, Antczak A, Pietras T, Bialasiewicz P. Verhoogd gehalte aan thiobarbituurzuur-reactieve stoffen en waterstofperoxide in het uitgeademde condensaat van patiënten met stabiele chronische obstructieve longziekte: geen significant effect van het roken van sigaretten. Adem Med. 1999;93:389�396.
120. Kelly FJ, Mudway IS. Eiwitoxidatie op het grensvlak tussen lucht en longen. Aminozuren. 2003;25:375�396.
121. Dean RT, Roberts CR, Jessup W. Fragmentatie van extracellulaire en intracellulaire polypeptiden door vrije radicalen. Prog Clin Biol Res. 1985;180:341�350.
122. Kek RG. Het gebruik van t-butylhydroperoxide als sonde voor methionine-oxidatie in eiwitten. Anale Biochemie. 1996;236:56�62.
123. Davies KJ. Eiwitbeschadiging en afbraak door zuurstofradicalen. I. Algemene aspecten. J Biol Chem. 1987;262:9895�9901.
124. Stadtman ER. Metaalion-gekatalyseerde oxidatie van eiwitten: biochemisch mechanisme en biologische gevolgen. Gratis Radic Biol Med.
1990; 9: 315.
125. Fucci L, Oliver CN, Coon MJ, Stadtman ER. Inactivering van belangrijke metabole enzymen door oxidatiereacties met gemengde functies: mogelijke implicaties voor eiwitomzet en veroudering. Proc Natl Acad Sci US A. 1983;80:1521-1525.
126. Stadtman ER, Moskovitz J, Levine RL. Oxidatie van methionineresiduen van eiwitten: biologische gevolgen. Antioxide Redox-signaal. 2003;5:577�582.
127. Stadtman ER, Levine RL. Door vrije radicalen gemedieerde oxidatie van vrije aminozuren en aminozuurresiduen in eiwitten. Aminozuren. 2003;25:207�218.
128. Stadtman ER. Eiwitoxidatie bij veroudering en ouderdomsziekten. Ann NY Acad Sci. 2001;928:22�38.
129. Shacter E. Kwantificering en betekenis van eiwitoxidatie in biologische monsters. Geneesmiddelen Metab Rev. 2000;32:307�326.
130. Poli G, Leonarduzzi G, Biasi F, Chiarpotto E. Oxidatieve stress en celsignalering. Curr Med Chem. 2004;11:1163�1182.
131. Neufeld G, Cohen T, Gengrinovitch S, Poltorak Z. Vasculaire endotheliale groeifactor (VEGF) en zijn receptoren. FASEB J. 1999;13:9�22.
132. Sundaresan M, Yu ZX, Ferrans VJ, Sulciner DJ, Gutkind JS, et al. Regulering van het genereren van reactieve zuurstofsoorten in fibroblasten door Rac1. Biochem J. 1996;318:379-382.
133. Zon T, Oberley LW. Redox-regulatie van transcriptionele activatoren. Gratis radicaal Biol Med. 1996;21:335�348.
134. Klatt P, Molina EP, De Lacoba MG, Padilla CA, Martinez-Galesteo E, Barcena JA, Lamas S. Redox-regulatie van c-Jun DNA-binding door reversibele S-glutathiolering. FASEB J. 1999;13:1481-1490.
135. Reynaert NL, Ckless K, Guala AS, Wouters EF, van der Vliet A, Janssen Heininger
JM. In situ detectie van S-glutathionyleerde eiwitten na glutaredoxine-1 gekatalyseerde cysteïnederivatisering. Biochim Biophys Acta. 2006;1760:380�387.
136. Reynaert NL, Wouters EF, Janssen-Heininger YM. Modulatie van glutaredoxin-1
expressie in een muismodel van allergische luchtwegaandoeningen. Am J Adem Cell Mol Biol. 2007;36:147�151.
137. Filomeni G, Rotilio G, Ciriolo MR. Celsignalering en het glutathion-redoxsysteem. Biochem Pharmacol. 2002;64:1057�1064.
138. Pande V, Ramos MJ. Moleculaire herkenning van 15-deoxydelta (12,14) prostaglandine J(2) door nucleaire factor-kappa B en andere cellulaire eiwitten. Bioorg Med Chem Lett. 2005;15:4057�4063.
139. Perkins ND. Integratie van celsignaleringsroutes met NF-kappaB- en IKK-functie. Nat Rev Mol Cel Biol. 2007;8:49�62.
140. Gilmore TD. Inleiding tot NF-kappaB: spelers, paden, perspectieven. Oncogeen. 2006;25:6680�6684.
141. Hirota K, Murata M, Sachi Y, Nakamura H, Takeuchi J, Mori K, Yodoi J. Verschillende rollen van thioredoxine in het cytoplasma en in de kern. Een tweestapsmechanisme van redoxregulatie van transcriptiefactor NF-kappaB. J Biol Chem. 1999;274:27891�27897.
142. Afdeling PA. De rol van complement, chemokines en regulerende cytokines bij acuut longletsel. Ann NY Acad Sci. 1996;796:104�112.
143. Akira S, Kishimoto A. NF-IL6 en NF-kB in cytokine-genregulatie. Adv Immunol. 1997;65:1�46.
144. Meyer M, Schreck R, Baeuerle PA. H2O2 en antioxidanten hebben tegengestelde effecten op de activering van NF-kappa B en AP-1 in intacte cellen: AP-1 als secundaire antioxidant-responsieve factor. EMBO J. 1993;12:2005-2015.
145. Abate C, Patel L, Rausher FJ, Curran T. Redox-regulatie van fos en jun DNA-bindende activiteit in vitro. Wetenschap. 1990;249:1157�1161.
146. Galter D, Mihm S, Droge W. Duidelijke effecten van glutathiondisulfide op de nucleaire transcriptiefactoren kB en het activator eiwit-1. Eur J Biochem. 1994;221:639�648.
147. Hirota K, Matsui M, Iwata S, Nishiyama A, Mori K, Yodoi J. AP-1-transcriptieactiviteit wordt gereguleerd door een directe associatie tussen thioredoxine en Ref-1. Proc Natl Acad Sci VS A. 1997;94: 3633�3638.
IFM's Find A Practitioner-tool is het grootste verwijzingsnetwerk in Functional Medicine, opgericht om patiënten te helpen bij het vinden van Functional Medicine-beoefenaars overal ter wereld. IFM Certified Practitioners worden als eerste vermeld in de zoekresultaten, gezien hun uitgebreide opleiding in functionele geneeskunde