Back Clinic Oxidatieve Stress Chiropractie en Functionele Geneeskunde Team. Oxidatieve stress wordt gedefinieerd als een verstoring in de balans tussen de productie van reactieve zuurstof (vrije radicalen) en de afweer van antioxidanten. Met andere woorden, het is een disbalans tussen de productie van vrije radicalen en het vermogen van het lichaam om de schadelijke effecten tegen te gaan of te ontgiften door neutralisatie door antioxidanten. Oxidatieve stress leidt tot veel pathofysiologische aandoeningen in het lichaam. Deze omvatten neurodegeneratieve ziekten, dwz de ziekte van Parkinson, de ziekte van Alzheimer, genmutaties, kankers, chronisch vermoeidheidssyndroom, fragiel X-syndroom, hart- en bloedvataandoeningen, atherosclerose, hartfalen, hartaanval en ontstekingsziekten. Oxidatie vindt plaats onder een aantal omstandigheden:
de cellen gebruiken glucose om energie te maken
het immuunsysteem bestrijdt bacteriën en veroorzaakt ontstekingen
de lichamen ontgiften verontreinigende stoffen, pesticiden en sigarettenrook
Er zijn miljoenen processen op elk moment in ons lichaam die kunnen leiden tot oxidatie. Hier zijn een paar symptomen:
Vermoeidheid
Geheugenverlies en of hersenmist
Spier- en / of gewrichtspijn
Rimpels samen met grijs haar
Verminderd gezichtsvermogen
Hoofdpijn en gevoeligheid voor lawaai
Gevoeligheid voor infecties
Het kiezen van biologisch voedsel en het vermijden van gifstoffen in uw omgeving maakt een groot verschil. Dit, samen met het verminderen van stress, kan gunstig zijn bij het verminderen van oxidatie.
Moe wakker worden, zelfs na zes of meer uur slaap?
Onder een hoge mate van stress?
Als u een van deze situaties ervaart, kan dit te wijten zijn aan uw melatonine- en cortisolspiegels die uw lichaam en circadiane ritme beïnvloeden.
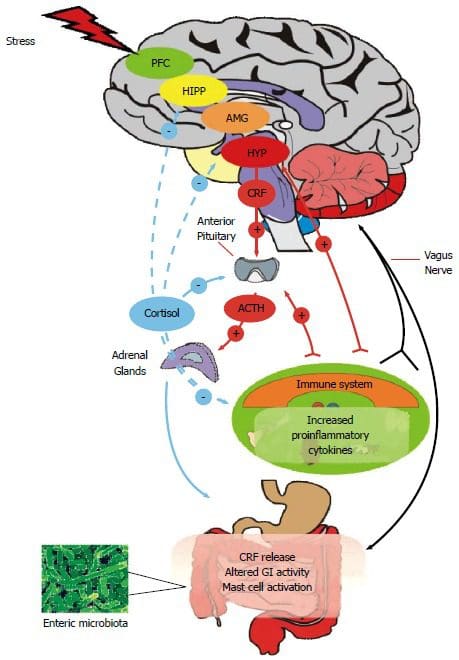
Over de hele wereld hebben miljoenen mensen moeite met slapen. In de Verenigde Staten zijn er ongeveer 50-70 miljoen mensen die een slechte slaapkwaliteit hebben. Wanneer een persoon minder dan acht uur heeft geslapen, worden ze moe en kunnen ze veel problemen krijgen, vooral als hun leven hectisch is. Met een hectische levensstijl en slecht slapen, kan het ervoor zorgen dat het lichaam weinig energie heeft om elke taak te klaren, het stresshormoon cortisol zal worden verhoogd en ziekten zoals hoge bloeddruk en diabetes problemen kunnen veroorzaken die chronisch kunnen zijn als dit niet het geval is. behandeld.
In de functionele endocrinologie zijn melatonine en cortisol hormonen die het lichaam van nature aanmaakt. Het cortisolhormoon of het stresshormoon helpt het lichaam in een staat van "vechten of vluchten" te komen, wat een goede zaak kan zijn voor iedereen die een project uitvoert of een sollicitatiegesprek heeft. Hoewel de cortisolhormoonspiegels hoog zijn, kan dit ertoe leiden dat het lichaam complicaties krijgt zoals ontstekingen, chronische oxidatieve stress en hoge bloeddruk.
Het circadiaans ritme van melatonine
Met het melatonine hormoon vertelt dit hormoon het lichaam wanneer het tijd is om te slapen. Soms hebben mensen echter moeite met slapen, en het nemen van melatoninesupplementen kan het lichaam juist ontspannen en de persoon dus in slaap laten vallen. Omdat de pijnappelklier melatonine uit de hersenen produceert, kan het ook in de ogen, het beenmerg en de darm worden gevonden om het lichaam te ontspannen en de persoon op natuurlijke wijze in slaap te laten vallen. Sommige onderzoeken tonen aan dat het circadiane ritme van de pijnappelklier die melatonine produceert. Door dit te doen, toont het onderzoek aan dat de toediening van melatonine kan:
One: slaap opwekken bij personen die moeite hebben met inslapen.
Twee: remt het lichaam om op natuurlijke wijze wakker te worden uit de circadiane pacemaker.
Drie: verschuif de circadiane biologische klokken om de slaapopname te verhogen wanneer een persoon op een vroeger tijdstip probeert te slapen om de volledige acht uur durende slaap te krijgen.
Wanneer iemand een baan van 9 tot 5 heeft, staan ze op met hun lichaam en ontspannen ze hun lichaam na een zware dag op het werk. Studies ontdekten dat de melatonine- en cortisolhormonen enorm helpen bij het reguleren van het 24-uurspatroon van de lichaamsfunctie en -reacties. Met de hormoonproductiecyclus van het lichaam kan het worden verstoord als de persoon 's avonds laat wakker blijft of overdag slaapt. Wanneer dit gebeurt, kan de persoon storende stoornissen krijgen zoals stemmingswisselingen, duizeligheid, prikkelbaar en depressief zijn en stofwisselingsstoornissen hebben. Niet alleen dat, maar ook het immuunsysteem en het endocriene systeem van het lichaam kunnen worden beschadigd, waardoor het lichaam een gastheer wordt voor infecties en ziekten.
Er zijn meer onderzoeken gedaan naar de circadiane ritmes in het lichaam, zoals: de studies tonen aan hoe mensen die in de nachtploeg werken in verband zijn gebracht met een groot aantal nadelige gezondheidsproblemen die het cardiovasculaire en gastro-intestinale systeem aantasten en het metabolische systeem verstoren. Iedereen die nachtploeg heeft gewerkt, moet zijn slaapschema wijzigen en zich aanpassen aan de snelle heroriëntatie in zijn slaap-/waakschema om naar zijn werk te gaan en zijn werk te doen. Aangezien iedereen met een ploegenschema werkt, kan dit stressvol zijn en de lichaamsprestaties van een werknemer beïnvloeden, evenals de melatonine- en cortisolsecretie beïnvloeden.
Manieren om cortisol en melatonine te ondersteunen
Verrassend genoeg zijn er manieren om de cortisolspiegel te verlagen en ervoor te zorgen dat de melatoninespiegels correct werken om het lichaam te laten functioneren. Om de cortisolspiegel te verlagen, moet een persoon meditatieve oefeningen doen, een plezierige hobby vinden en, belangrijker nog, diepe ademhalingsoefeningen proberen om het lichaam te ontspannen van ongewenste stress. Met diepe ademhalingsoefeningen kan het het lichaam helpen alle spanning die een persoon vasthoudt los te laten, en de spieren in het lichaam beginnen te ontspannen en het bloed begint te stromen. Met de melatoninespiegels werken ze samen met het circadiane ritme van het lichaam en zorgen ervoor dat het lichaam weet wanneer het tijd is om wakker te worden, te slapen en te eten. Het melatonine-hormoon kan ook helpen bij het reguleren van de lichaamstemperatuur, bloeddruken hormoonspiegels om ervoor te zorgen dat het goed functioneert. Wanneer er hoge niveaus van deze systemen zijn, kan dit ertoe leiden dat het lichaam chronische ziekten ontwikkelt en het lichaam daarbij schade toebrengt.
Onderzoek toont aan dat melatoninehormonen zich kunnen binden aan neurologische receptoren in het lichaam, waardoor ontspanning wordt bevorderd. Omdat melatonine zich bindt aan neurologische receptoren, kan het ook de zenuwactiviteit en het dopaminegehalte verminderen om de ogen zwaar te maken, waardoor de persoon in slaap valt.
Conclusie
Omdat het lichaam op natuurlijke wijze melatonine- en cortisolspiegels kan produceren om ervoor te zorgen dat het lichaam de hele dag niet overmatig gestrest raakt. Omdat melatonine samenwerkt met het circadiane ritme van het lichaam, weet het lichaam wanneer het op moet blijven en in slaap moet vallen. Omdat iedereen een hectisch schema heeft, is het essentieel om de tijd te nemen en te ontspannen en een gezond slaapschema te volgen, zodat het lichaam gezond kan zijn en kan functioneren. Sommige producten zijn hier om ervoor te zorgen dat het endocriene systeem goed functioneert en de bijnieren en het suikermetabolisme ondersteunt.
De reikwijdte van onze informatie is beperkt tot chiropractie, musculoskeletale en nerveuze gezondheidsproblemen of artikelen, onderwerpen en discussies over functionele geneeskunde. We gebruiken functionele gezondheidsprotocollen om verwondingen of aandoeningen van het bewegingsapparaat te behandelen. Ons kantoor heeft een redelijke poging gedaan om ondersteunende citaten te verstrekken en heeft de relevante onderzoeksstudie of studies geïdentificeerd die onze functies ondersteunen. We maken op verzoek ook kopieën van ondersteunende onderzoeksstudies beschikbaar voor het bestuur en / of het publiek. Als u het onderwerp hierboven verder wilt bespreken, kunt u Dr. Alex Jimenez vragen of contact met ons opnemen via 915-850-0900.
Referenties:
Cajochen, C, et al. "De rol van melatonine in de regulatie van menselijke circadiaanse ritmes en slaap." Journal of Neuroendocrinology, US National Library of Medicine, april 2003, www.ncbi.nlm.nih.gov/pubmed/12622846.
James, Francine O, et al. �Circadische ritmes van melatonine, cortisol en klokgenexpressie tijdens gesimuleerde nachtploeg.� Slaap, Associated Professional Sleep Societies, LLC, november 2007, www.ncbi.nlm.nih.gov/pmc/articles/PMC2082093/.
Monteleone, P, et al. �Tijdelijke relatie tussen melatonine en Cortisol-reacties op nachtelijke fysieke stress bij mensen.� Psychoneuroendocrinology, US National Library of Medicine, 1992, www.ncbi.nlm.nih.gov/pubmed/1609019.
Raman, Ryan. �Hoe melatonine u kan helpen slapen en u beter te voelen.� Healthline, Healthline Media, 3 september 2017, www.healthline.com/nutrition/melatonine-and-sleep.
Zamanian, Zahra, et al. �Overzicht van veranderingen in Cortisol en melatonine circadiane ritmes in de bewakers van de Shiraz University of Medical Sciences.� International Journal of Preventive Medicine, Medknow Publications & Media Pvt Ltd, juli 2013, www.ncbi.nlm.nih.gov/pmc/articles/PMC3775223/.
Moderne integratieve en functionele geneeskunde- Esse Quam Videri
Door individuen te informeren over hoe de National University of Health Sciences kennis verschaft aan toekomstige generaties die een verschil willen maken in de wereld. De universiteit biedt een breed scala aan medische beroepen voor functionele en integratieve geneeskunde.
Heb je je ooit afgevraagd waarom je je traag voelt van een lange dag? Of voel je je misselijk als je iets slechts hebt gegeten of teveel hebt gegeten van je favoriete eten? Zou het kunnen dat je darmen tekenen van stress en ongemak vertonen door bepaalde gewoonten die je misschien tegenkomt en waarvan je niet eens wist?
In ons vorige artikel hadden we het over de zes soorten voedsel dat onze darm gezond moet zijn. Sinds onze onderbuik bevat biljoenen microbiomen, zowel goede als slechte, deze microbiomen spelen een belangrijke rol in onze algehele gezondheid. Een gezond microbioom verbetert onze uw darmgezondheid, gezondheid van het hart, hersengezondheid, controleert ons gewicht en regelt onze bloedsuikerspiegel. Met de goede bacteriën in onze darm, profiteren de bacteriën ons van een goed spijsverteringsstelsel en vernietigen ze de schadelijke bacteriën. Maar bepaalde levensstijlen en voedingskeuzes kunnen de slechte bacteriën juist verhogen en de goede bacteriën en de algehele gezondheid verlagen.
Hier zijn vijf verrassende levensstijlkeuzes die je darmen kwetsen:
Geen breed scala aan voedingsmiddelen eten
Onze darmen spelen een belangrijke rol in onze algehele gezondheid. Als we goed, heel voedsel eten, zijn onze darmen gelukkiger; we hebben meer energie om elke taak te voltooien die ons wordt toegeworpen en we krijgen voedingsstoffen voor onze darmflora. De afgelopen decennia leunen we echter meer op bewerkte voedingsmiddelen vanwege de economische druk van verhoogde voedselproducties. FOA verklaarde dat '75 procent van het voedsel in de wereld wordt gegenereerd door slechts 12 planten en vijf diersoorten' en dat is erg slecht voor onze darmflora.
Hier bij Injury Medical & Chiropractic Clinic informeren we onze patiënten over het belang van het eten van voedzaam, volwaardig voedsel om niet alleen een gezonde darm, maar ook een gezonde geest te bevorderen. Wanneer het lichaam kennismaakt met een grote verscheidenheid aan hele voedingsmiddelen (met een hoog vezelgehalte), begint onze darm de schade te herstellen van bewerkte voedingsmiddelen die we mogelijk intern hebben geconsumeerd.
Als u prebiotica echter buiten beschouwing laat in uw dieet, bent u: schadelijk voor uw spijsvertering. Zonder prebiotica vertraagt ons spijsverteringssysteem de ontwikkeling en diversiteit van onze darmflora. Dus om een gezond microbioom ontwikkeling, moet u voedingsmiddelen met zowel verteerbare als onverteerbare vezels in uw dieet opnemen. Sommige voedingsmiddelen in deze categorie zijn haver, noten, uien, knoflook, prei, asperges, bananen, peren, kikkererwten en bonen.
Vasthouden aan een vezelrijk dieet is misschien een uitdaging, maar er is de mogelijkheid om prebiotische supplementen te nemen. Als u een voedselallergeen of voedselovergevoeligheid heeft voor voedingsmiddelen met een hoog vezelgehalte, neem dan: prebiotische supplementen kan daadwerkelijk helpen Bifidobacterium en Faecalibacterium in uw darm te laten groeien en gunstig zijn voor uw gezondheid zonder het ongemak.
Overmatig alcoholgebruik
Elke volwassene drinkt wel eens alcohol. Ja, het is een van die dranken die je helpen een beetje te ontspannen na een lange dag, maar te veel ervan kan leiden tot alcoholmisbruik en verslaving. Dus, wist je dat het consumeren van zoveel alcohol slecht is voor? je hart, lever en hersenen; waardoor uw darmgezondheid wordt geschaad en u dysbiose krijgt?
Eén studie verklaarde dat de alcoholisten met dysbiose een lagere mediane overvloed aan Bacteroidetes en een hoge overvloed aan Proteobacteria hadden. Degenen die geen alcoholisten waren, werden niet beïnvloed door het onderzoek.
Echter; er is goed nieuws over jezelf beperken tot alcoholisme en dat het gunstig kan zijn voor je darmbacteriën. Als u matig rode wijn op een verantwoorde manier consumeert, polyfenolen in de wijn kan je darmflora ten goede komen. Dus geniet af en toe van een glas wijn als een kleine traktatie die niet als vanzelfsprekend mag worden beschouwd.
Onvoldoende slaap
In een van de vorige artikelen hebben we het gehad over het bereiken van een goede nachtrust door kruiden. Wanneer we weinig tot geen slaap krijgen door ons hectische leven, beïnvloedt dit ons door verschillende gezondheidsproblemen, waaronder: hartziekte en zwaarlijvigheid. In een 2016 studie, ontdekten onderzoekers na twee dagen het effect van kortdurend slaaptekort op de darmflora.
Wanneer ons lichaam niet de aanbevolen 8 uur slaap krijgt, eist onze darm een enorme tol omdat we ons traag en uitgeput voelen. Dus, om ervoor te zorgen dat onze darm microbioom zal worden geregeld, raden we aan om uw elektronische apparaten ten minste 30 minuten voordat u zich klaarmaakt om te settelen voor de nacht uit te schakelen. Doe alle lichten uit en drink geen vloeistoffen ten minste twee uur voor het slapengaan, sluit je ogen en haal diep adem in een meditatieve staat, en ontspan terwijl je wegdrijft in de slaapstad.
Onvoldoende lichaamsbeweging
Door onze snelle levensstijl en stressvolle banen is het moeilijk om tijd te vinden om te sporten. Maar als we echt tijd vinden om te sporten, voelen we ons niet alleen goed; maar ons lichaam en onze darmen voelen ook goed aan. Er komen echter altijd dingen naar voren als we een trainingsroutine hebben en we het sporten helemaal moeten overslaan. Het overkomt ons allemaal en het is moeilijk om verder te gaan waar we waren gebleven toen we probeerden te oefenen.
Als we minstens een paar keer per week niet sporten, eist ons lichaam een enorme tol van ons naarmate we aankomen, onze stress is veel te hoogen we hebben een grotere kans van het krijgen van een chronische ziekte. Wanneer dit gebeurt, is onze darmflora een enorm nadeel. Hier in de kliniek streven we ernaar onze patiënten te informeren over het belang van lichaamsbeweging en dat het niet alleen hun leven verandert, maar ook hun stemming volledig.
Ga echter niet zomaar in een zware trainingsroutine waarbij u uzelf zult verwonden. Begin met een training met een lage intensiteit en bouw deze vervolgens op terwijl je bezig bent, want je darmflora zal je er dankbaar voor zijn.
Als laatste woord willen we hier bij Injury Medical u op de hoogte houden over voeding en manieren om u te helpen uw kwalen te verbeteren met deze 5 verrassingen. Maar om u ook voor te lichten over wat uw darmen kan kwetsen. Met deze verrassingen en kleine veranderingen in je dagelijkse leven, zullen je darmen je bedanken voor de lange termijn.
NCBI-bronnen
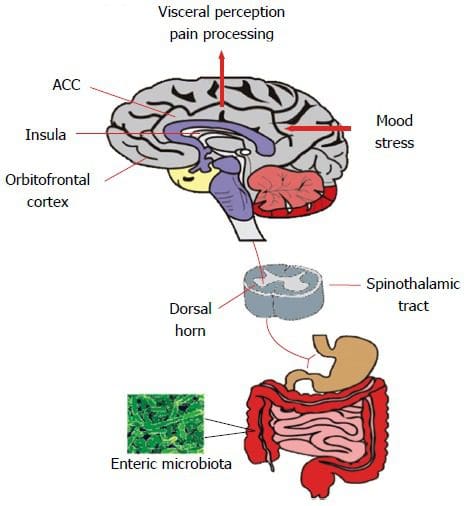
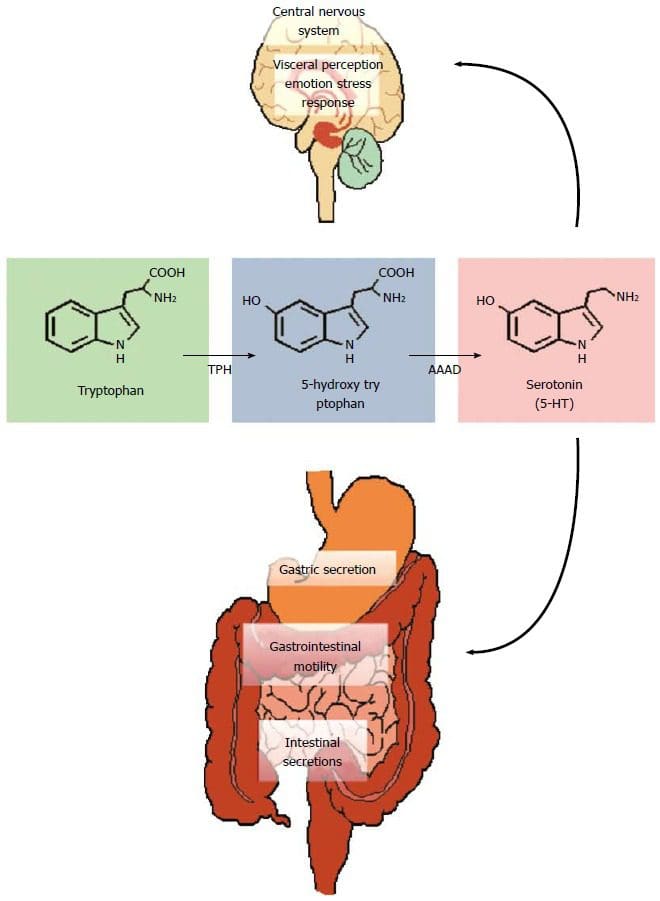
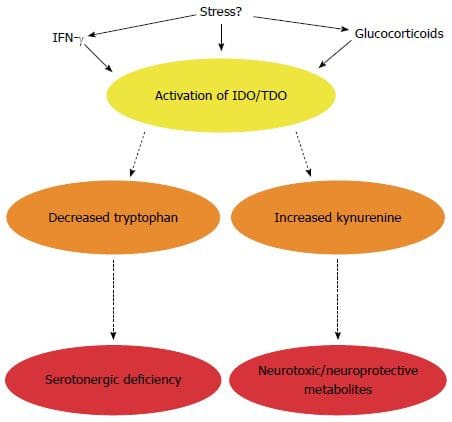
Volgens bewijs uit een onderzoeksstudie uit 2016, is het immuunsysteem van de darm van fundamenteel belang voor het voorkomen van een verscheidenheid aan ziekten en kan het vaak bijdragen aan stofwisselingsstoornissen. Het kan echter ook helpen om een behandeldoel te stellen bij het observeren van systemische ontstekingen bij insulineresistentie. Bovendien is gemodificeerde darmimmuniteit in verband gebracht met veranderingen in de darmmicrobiota, darmbarrièrefunctie, in de darm verblijvende immuuncellen en resistentie tegen antigenen die het gastro-intestinale of GI-systeem binnendringen. Hoewel eerder werd aangenomen dat dit het gevaar van slokdarmaandoeningen verhoogt, waaronder pathogene infecties en chronische ontstekingen, die uiteindelijk kunnen leiden tot chronische gezondheidsproblemen.
Ketonlichamen worden aangemaakt door de lever en gebruikt als energiebron wanneer glucose niet direct beschikbaar is in het menselijk lichaam. De twee belangrijkste ketonlichamen zijn acetoacetaat (AcAc) en 3-beta-hydroxybutyraat (3HB), terwijl aceton het derde en minst voorkomende ketonlichaam is. Ketonen zijn altijd aanwezig in het bloed en hun niveaus stijgen tijdens vasten en langdurige inspanning.�ketogenese is het biochemische proces waarbij organismen ketonlichamen produceren door de afbraak van vetzuren en ketogene aminozuren.
Ketonlichamen worden voornamelijk gegenereerd in de mitochondriën van levercellen. Ketogenese treedt op wanneer er lage glucosespiegels in het bloed zijn, vooral nadat andere cellulaire koolhydraatvoorraden, zoals glycogeen, zijn uitgeput. Dit mechanisme kan ook optreden als er onvoldoende insuline is. De productie van ketonlichamen wordt uiteindelijk gestart om energie beschikbaar te maken die in het menselijk lichaam wordt opgeslagen als vetzuren. Ketogenese vindt plaats in de mitochondriën waar het onafhankelijk wordt gereguleerd.
Abstract
Het ketonlichaammetabolisme is een centraal knooppunt in fysiologische homeostase. In deze review bespreken we hoe ketonen een discrete fijnafstemming van metabolische rollen hebben die de prestaties van organen en organismen optimaliseren in verschillende nutriëntenresten en beschermen tegen ontstekingen en verwondingen in meerdere orgaansystemen. Traditioneel gezien als metabolische substraten die alleen worden gebruikt bij koolhydraatbeperking, onderstrepen recente waarnemingen het belang van ketonlichamen als vitale metabole en signaalmediatoren wanneer koolhydraten in overvloed aanwezig zijn. Als aanvulling op een repertoire van bekende therapeutische opties voor ziekten van het zenuwstelsel, zijn prospectieve rollen voor ketonlichamen bij kanker ontstaan, evenals intrigerende beschermende rollen in hart en lever, waardoor therapeutische opties ontstaan bij obesitas-gerelateerde en cardiovasculaire aandoeningen. Controverses in ketonmetabolisme en -signalering worden besproken om klassieke dogma's te verzoenen met hedendaagse observaties.
Introductie
Ketonlichamen zijn een essentiële alternatieve metabolische brandstofbron voor alle domeinen van het leven, eukarya, bacteriën en archaea (Aneja et al., 2002; Cahill GF Jr, 2006; Krishnakumar et al., 2008). Het ketonlichaammetabolisme bij mensen is gebruikt om de hersenen van brandstof te voorzien tijdens episodische perioden van tekort aan voedingsstoffen. Ketonlichamen zijn verweven met cruciale metabole routes bij zoogdieren zoals ?-oxidatie (FAO), de tricarbonzuurcyclus (TCA), gluconeogenese, de novo lipogenese (DNL) en biosynthese van sterolen. Bij zoogdieren worden ketonlichamen voornamelijk in de lever geproduceerd uit FAO-afgeleid acetyl-CoA, en ze worden getransporteerd naar extrahepatische weefsels voor terminale oxidatie. Deze fysiologie biedt een alternatieve brandstof die wordt versterkt door relatief korte perioden van vasten, waardoor de beschikbaarheid van vetzuren toeneemt en de beschikbaarheid van koolhydraten afneemt (Cahill GF Jr, 2006; McGarry en Foster, 1980; Robinson en Williamson, 1980). De oxidatie van ketonen levert een belangrijke bijdrage aan het algehele energiemetabolisme van zoogdieren in extrahepatische weefsels in een groot aantal fysiologische toestanden, waaronder vasten, uithongering, de neonatale periode, na inspanning, zwangerschap en het volgen van koolhydraatarme diëten. De circulerende totale ketonlichaamconcentraties bij gezonde volwassen mensen vertonen normaal gesproken circadiane schommelingen tussen ongeveer 100-250 �M, stijgen tot ~1 mM na langdurige inspanning of 24 uur vasten, en kunnen accumuleren tot wel 20 mM in pathologische toestanden zoals diabetische ketoacidose ( Cahill GF Jr, 2006; Johnson et al., 1969b; Koeslag et al., 1980; Robinson en Williamson, 1980; Wildenhoff et al., 1974). De menselijke lever produceert tot 300 g ketonlichamen per dag (Balasse en Fery, 1989), die tussen de 5% van het totale energieverbruik bijdragen in gevoede, nuchtere en uitgehongerde toestanden (Balasse et al., 20; Cox et al. al., 1978).
Recente studies benadrukken nu de dwingende rol voor ketonlichamen in het celmetabolisme, homeostase en signalering van zoogdieren onder een grote verscheidenheid aan fysiologische en pathologische toestanden. Afgezien van het dienen als energiebrandstof voor extrahepatische weefsels zoals hersenen, hart of skeletspieren, spelen ketonlichamen een cruciale rol als signaalbemiddelaars, aanjagers van post-translationele eiwitmodificatie (PTM) en modulatoren van ontsteking en oxidatieve stress. In deze review bieden we zowel klassieke als moderne opvattingen over de pleiotrope rollen van ketonlichamen en hun metabolisme.
Overzicht van ketonlichaammetabolisme
De snelheid van hepatische ketogenese wordt bepaald door een georkestreerde reeks fysiologische en biochemische transformaties van vet. Primaire regulatoren omvatten lipolyse van vetzuren uit triacylglycerolen, transport naar en door het hepatocytplasmamembraan, transport naar mitochondriën via carnitinepalmitoyltransferase 1 (CPT1), de ?-oxidatiespiraal, TCA-cyclusactiviteit en tussenliggende concentraties, redoxpotentiaal en de hormonale regulatoren van deze processen, voornamelijk glucagon en insuline [besproken in (Arias et al., 1995; Ayte et al., 1993; Ehara et al., 2015; Ferre et al., 1983; Kahn et al., 2005; McGarry en Foster , 1980; Williamson et al., 1969)]. Klassiek wordt ketogenese gezien als een overlooproute, waarbij van ?-oxidatie afgeleid acetyl-CoA de citraatsynthaseactiviteit en/of de beschikbaarheid van oxaalacetaat voor condensatie om citraat te vormen, overschrijdt. Tussenproducten met drie koolstofatomen vertonen anti-ketogene activiteit, vermoedelijk vanwege hun vermogen om de oxaalacetaatvoorraad voor acetyl-CoA-consumptie uit te breiden, maar de acetyl-CoA-concentratie in de lever alleen bepaalt niet de ketogene snelheid (Foster, 1967; Rawat en Menahan, 1975; Williamson et al., 1969). De regulatie van ketogenese door hormonale, transcriptionele en post-translationele gebeurtenissen ondersteunen samen het idee dat de moleculaire mechanismen die de ketogene snelheid verfijnen, onvolledig worden begrepen (zie Regulatie van HMGCS2 en SCOT/OXCT1).
Ketogenese vindt voornamelijk plaats in de mitochondriale matrix van de lever met snelheden die evenredig zijn met de totale vetoxidatie. Na transport van acylketens over de mitochondriale membranen en ?-oxidatie, katalyseert de mitochondriale isovorm van 3-hydroxymethylglutaryl-CoA-synthase (HMGCS2) het lot waarbij condensatie van acetoacetyl-CoA (AcAc-CoA) en acetyl-CoA wordt bewerkstelligd om HMG-CoA te genereren (Fig. 1A). HMG-CoA-lyase (HMGCL) splitst HMG-CoA om acetyl-CoA en acetoacetaat (AcAc) vrij te maken, en de laatste wordt gereduceerd tot d-?-hydroxybutyraat (d-?OHB) door fosfatidylcholine-afhankelijke mitochondriale d-?OHB-dehydrogenase ( BDH1) in een NAD+/NADH-gekoppelde bijna-evenwichtsreactie (Bock en Fleischer, 1975; LEHNINGER et al., 1960). De BDH1-evenwichtsconstante bevordert de d-?OHB-productie, maar de verhouding van AcAc/d-?OHB-ketonlichamen is recht evenredig met de mitochondriale NAD+/NADH-verhouding, en dus moduleert BDH1-oxidoreductase-activiteit de mitochondriale redoxpotentiaal (Krebs et al., 1969; Williamson et al., 1967). AcAc kan ook spontaan decarboxyleren tot aceton (Pedersen, 1929), de bron van zoete geur bij mensen die lijden aan ketoacidose (dwz totale serumketonlichamen > ~7 mM; AcAc pKa 3.6, ?OHB pKa 4.7). De mechanismen waardoor ketonlichamen door het mitochondriale binnenmembraan worden getransporteerd, zijn niet bekend, maar AcAc/d-?OHB worden door cellen vrijgemaakt via monocarboxylaattransporters (bij zoogdieren, MCT 1 en 2, ook bekend als opgeloste stofdrager 16A-familieleden 1 en 7) en in de bloedsomloop getransporteerd naar extrahepatische weefsels voor terminale oxidatie (Cotter et al., 2011; Halestrap en Wilson, 2012; Halestrap, 2012; Hugo et al., 2012). Concentraties van circulerende ketonlichamen zijn hoger dan die in de extrahepatische weefsels (Harrison en Long, 1940), wat aangeeft dat ketonlichamen langs een concentratiegradiënt worden getransporteerd. Verlies van functie-mutaties in MCT1 zijn geassocieerd met spontane aanvallen van ketoacidose, wat een cruciale rol suggereert bij de import van ketonlichamen.
Met uitzondering van mogelijke omleiding van ketonlichamen naar niet-oxidatieve lotsbestemmingen (zie Niet-oxidatieve metabolische lotgevallen van ketonlichamen), hebben hepatocyten niet het vermogen om de ketonlichamen die ze produceren te metaboliseren. Ketonlichamen die de novo door de lever worden gesynthetiseerd, worden (i) in mitochondriën van extrahepatische weefsels gekataboliseerd tot acetyl-CoA, dat beschikbaar is voor de TCA-cyclus voor terminale oxidatie (Fig. 1A), (ii) omgeleid naar de lipogenese- of sterolsyntheseroutes ( Fig. 1B), of (iii) uitgescheiden in de urine. Als alternatieve energetische brandstof worden ketonlichamen gretig geoxideerd in hart, skeletspieren en hersenen (Balasse en Fery, 1989; Bentourkia et al., 2009; Owen et al., 1967; Reichard et al., 1974; Sultan, 1988 ). Extrahepatische mitochondriale BDH1 katalyseert de eerste reactie van ?OHB-oxidatie en zet deze om in AcAc (LEHNINGER et al., 1960; Sandermann et al., 1986). Een cytoplasmatisch d-?OHB-dehydrogenase (BDH2) met slechts 20% sequentie-identiteit met BDH1 heeft een hoge Km voor ketonlichamen en speelt ook een rol bij de ijzerhomeostase (Davuluri et al., 2016; Guo et al., 2006) . In extrahepatische mitochondriale matrix wordt AcAc geactiveerd tot AcAc-CoA door uitwisseling van een CoA-deel van succinyl-CoA in een reactie die wordt gekatalyseerd door een uniek zoogdier-CoA-transferase, succinyl-CoA:3-oxoacid-CoA-transferase (SCOT, CoA-transferase; gecodeerd door OXCT1), via een bijna-evenwichtsreactie. De vrije energie die vrijkomt door hydrolyse van AcAc-CoA is groter dan die van succinyl-CoA, wat de vorming van AcAc bevordert. Dus de oxidatieve flux van het ketonlichaam treedt op als gevolg van massale actie: een overvloedige toevoer van AcAc en snelle consumptie van acetyl-CoA via citraatsynthase bevordert AcAc-CoA (+ succinaat) vorming door SCOT. Met name, in tegenstelling tot glucose (hexokinase) en vetzuren (acyl-CoA-synthetasen), vereist de activering van ketonlichamen (SCOT) in een oxideerbare vorm geen investering van ATP. Een omkeerbare AcAc-CoA-thiolasereactie [gekatalyseerd door een van de vier mitochondriale thiolasen die worden gecodeerd door ACAA2 (codering van een enzym dat bekend staat als T1 of CT), ACAT1 (codering van T2), HADHA of HADHB] levert twee moleculen acetyl-CoA op, die de TCA-cyclus binnengaan (Hersh en Jencks, 1967; Stern et al., 1956; Williamson et al., 1971). Tijdens ketotische toestanden (dwz totale serumketonen > 500 �M), leveren ketonlichamen een belangrijke bijdrage aan het energieverbruik en worden ze snel in weefsels gebruikt totdat de oxidatie wordt opgenomen of verzadigt (Balasse et al., 1978; Balasse en Fery, 1989 Edmond et al., 1987). Een zeer klein deel van de van de lever afgeleide ketonlichamen kan gemakkelijk in de urine worden gemeten, en het gebruik en de reabsorptie door de nieren zijn evenredig met de circulerende concentratie (Goldstein, 1987; Robinson en Williamson, 1980). Tijdens zeer ketotische toestanden (> 1 mM in plasma) dient ketonurie als een semi-kwantitatieve reporter van ketose, hoewel de meeste klinische tests van ketonlichamen in de urine AcAc detecteren maar niet ?OHB (Klocker et al., 2013).
Ketogene substraten en hun impact op het hepatocytmetabolisme
Ketogene substraten omvatten vetzuren en aminozuren (Fig. 1B). Het katabolisme van aminozuren, in het bijzonder leucine, genereert ongeveer 4% van de ketonlichamen in post-absorptieve toestand (Thomas et al., 1982). Dus de acetyl-CoA-substraatpool om ketonlichamen te genereren, is voornamelijk afkomstig van vetzuren, omdat pyruvaat tijdens toestanden van verminderde koolhydraattoevoer de hepatische TCA-cyclus voornamelijk binnenkomt via anaplerose, dwz ATP-afhankelijke carboxylatie tot oxaalacetaat (OAA), of tot malaat (MAL), en niet oxidatieve decarboxylatie tot acetyl-CoA (Jeoung et al., 2012; Magnusson et al., 1991; Merritt et al., 2011). In de lever dragen glucose en pyruvaat verwaarloosbaar bij aan ketogenese, zelfs wanneer de decarboxylatie van pyruvaat tot acetyl-CoA maximaal is (Jeoung et al., 2012).
Acetyl-CoA vervult verschillende rollen die integraal deel uitmaken van het intermediaire metabolisme in de lever voorbij ATP-generatie via terminale oxidatie (zie ook De integratie van ketonlichaammetabolisme, post-translationele modificatie en celfysiologie). Acetyl-CoA activeert allosterisch (i) pyruvaatcarboxylase (PC), waardoor een metabool controlemechanisme wordt geactiveerd dat de anaplerotische toegang van metabolieten in de TCA-cyclus verhoogt (Owen et al., 2002; Scrutton en Utter, 1967) en (ii) pyruvaatdehydrogenase kinase, dat pyruvaatdehydrogenase (PDH) fosforyleert en remt (Cooper et al., 1975), waardoor de stroom van pyruvaat in de TCA-cyclus via anaplerose verder wordt verbeterd. Bovendien remt cytoplasmatisch acetyl-CoA, waarvan de pool wordt aangevuld met mechanismen die mitochondriaal acetyl-CoA omzetten in transporteerbare metabolieten, de oxidatie van vetzuren: acetyl-CoA-carboxylase (ACC) katalyseert de omzetting van acetyl-CoA in malonyl-CoA, het lipogene substraat en allosterische remmer van mitochondriaal CPT1 [besproken in (Kahn et al., 2005; McGarry en Foster, 1980)]. Dus de mitochondriale acetyl-CoA-pool reguleert en wordt gereguleerd door de overlooproute van ketogenese, die de belangrijkste aspecten van het intermediaire levermetabolisme orkestreert.
Niet-oxidatief metabolisch lot van ketonlichamen
Het overheersende lot van van de lever afgeleide ketonen is SCOT-afhankelijke extrahepatische oxidatie. AcAc kan echter worden geëxporteerd vanuit mitochondriën en worden gebruikt in anabole routes via conversie naar AcAc-CoA door een ATP-afhankelijke reactie die wordt gekatalyseerd door cytoplasmatische acetoacetyl-CoA-synthetase (AACS, figuur 1B). Deze route is actief tijdens de ontwikkeling van de hersenen en in de melkgevende borstklier (Morris, 2005; Robinson en Williamson, 1978; Ohgami et al., 2003). AACS wordt ook sterk tot expressie gebracht in vetweefsel en geactiveerde osteoclasten (Aguilo et al., 2010; Yamasaki et al., 2016). Cytoplasmatisch AcAc-CoA kan ofwel door cytosolisch HMGCS1 worden gericht op de biosynthese van sterol, of door een van de twee cytoplasmatische thiolasen worden gesplitst tot acetyl-CoA (ACAA1 en ACAT2), gecarboxyleerd tot malonyl-CoA, en bijdragen aan de synthese van vetzuren (Bergstrom et al. al., 1984; Edmond, 1974; Endemann et al., 1982; Geelen et al., 1983; Webber en Edmond, 1977).
Hoewel de fysiologische betekenis nog moet worden vastgesteld, kunnen ketonen zelfs in de lever als anabole substraten dienen. In kunstmatige experimentele contexten kan AcAc bijdragen tot wel de helft van het nieuw gesynthetiseerde lipide en tot 75% van het nieuwe gesynthetiseerde cholesterol (Endemann et al., 1982; Geelen et al., 1983; Freed et al., 1988). Omdat AcAc is afgeleid van onvolledige hepatische vetoxidatie, zou het vermogen van AcAc om bij te dragen aan lipogenese in vivo een futiele hepatische cyclus impliceren, waarbij van vet afgeleide ketonen kunnen worden gebruikt voor de productie van lipiden, een begrip waarvan de fysiologische betekenis experimentele validatie vereist, maar zou kunnen dienen adaptieve of onaangepaste rollen (Solinas et al., 2015). AcAc levert gretig cholesterogenese, met een lage AACS Km-AcAc (~ 50 �M) die AcAc-activering bevordert, zelfs in gevoede toestand (Bergstrom et al., 1984). De dynamische rol van cytoplasmatisch ketonmetabolisme is gesuggereerd in primaire embryonale neuronen van muizen en in van 3T3-L1 afgeleide adipocyten, aangezien AACS-knockdown de differentiatie van elk celtype verslechterde (Hasegawa et al., 2012a; Hasegawa et al., 2012b). Knockdown van AACS bij muizen in vivo verlaagde serumcholesterol (Hasegawa et al., 2012c). SREBP-2, een meestertranscriptieregulator van cholesterolbiosynthese, en peroxisoom proliferator geactiveerde receptor (PPAR)-? zijn AACS-transcriptieactivators en reguleren de transcriptie ervan tijdens de ontwikkeling van neurieten en in de lever (Aguilo et al., 2010; Hasegawa et al., 2012c). Alles bij elkaar genomen kan het metabolisme van het cytoplasmatische ketonlichaam belangrijk zijn bij bepaalde aandoeningen of natuurlijke ziektegeschiedenissen, maar het is niet toereikend om van de lever afgeleide ketonlichamen af te voeren, aangezien massale hyperketoniemie optreedt in de setting van selectieve verslechtering van het primaire oxidatieve lot via verlies van functiemutaties naar SCOT (Berry et al., 2001; Cotter et al., 2011).
Regulering van HMGCS2 en SCOT/OXCT1
De divergentie van een mitochondriaal van het gen dat codeert voor cytosolische HMGCS vond plaats vroeg in de evolutie van gewervelde dieren vanwege de noodzaak om hepatische ketogenese te ondersteunen bij soorten met hogere verhoudingen tussen hersenen en lichaamsgewicht (Boukaftane et al., 1994; Cunnane en Crawford, 2003). Natuurlijk voorkomende functieverlies HMGCS2-mutaties bij mensen veroorzaken aanvallen van hypoketotische hypoglykemie (Pitt et al., 2015; Thompson et al., 1997). Robuuste HMGCS2-expressie is beperkt tot hepatocyten en colonepitheel, en de expressie en enzymatische activiteit ervan worden gecoördineerd via diverse mechanismen (Mascaro et al., 1995; McGarry en Foster, 1980; Robinson en Williamson, 1980). Hoewel de volledige reikwijdte van fysiologische toestanden die HMGCS2 beïnvloeden verdere opheldering vereist, wordt de expressie en/of activiteit ervan gereguleerd tijdens de vroege postnatale periode, veroudering, diabetes, uithongering of inname van ketogeen dieet (Balasse en Fery, 1989; Cahill GF Jr, 2006 ; Girard et al., 1992; Hegardt, 1999; Satapati et al., 2012; Sengupta et al., 2010). Bij de foetus correleert de methylering van het 5�-flankerende gebied van het Hmgcs2-gen omgekeerd met de transcriptie ervan en wordt na de geboorte gedeeltelijk omgekeerd (Arias et al., 1995; Ayte et al., 1993; Ehara et al., 2015; Ferre et al. ., 1983). Evenzo vertoont hepatische Bdh1 een ontwikkelingsexpressiepatroon, dat toeneemt vanaf de geboorte tot het spenen, en wordt ook geïnduceerd door een ketogeen dieet op een fibroblastgroeifactor (FGF)-21-afhankelijke manier (Badman et al., 2007; Zhang et al., 1989 ). Ketogenese bij zoogdieren reageert zeer goed op zowel insuline als glucagon, en wordt respectievelijk onderdrukt en gestimuleerd (McGarry en Foster, 1977). Insuline onderdrukt de lipolyse van vetweefsel, waardoor de ketogenese van zijn substraat wordt beroofd, terwijl glucagon de ketogene flux verhoogt door een direct effect op de lever (Hegardt, 1999). Hmgcs2-transcriptie wordt gestimuleerd door forkhead-transcriptiefactor FOXA2, die wordt geremd via insuline-fosfatidylinositol-3-kinase/Akt, en wordt geïnduceerd door glucagon-cAMP-p300-signalering (Arias et al., 1995; Hegardt, 1999; Quant et al. , 1990; Thumelin et al., 1993; von Meyenn et al., 2013; Wolfrum et al., 2004; Wolfrum et al., 2003). PPAR? (Rodriguez et al., 1994) samen met zijn doelwit, FGF21 (Badman et al., 2007) induceren ook Hmgcs2-transcriptie in de lever tijdens uithongering of toediening van een ketogeen dieet (Badman et al., 2007; Inagaki et al., 2007 ). Inductie van PPAR? kan optreden vóór de overgang van foetale naar neonatale fysiologie, terwijl FGF21-activering in de vroege neonatale periode kan worden bevorderd via ?OHB-gemedieerde remming van histondeacetylase (HDAC)-3 (Rando et al., 2016). mTORC1 (zoogdierdoelwit van rapamycinecomplex 1) afhankelijke remming van PPAR? transcriptionele activiteit is ook een belangrijke regulator van Hmgcs2-genexpressie (Sengupta et al., 2010), en lever PER2, een meester-circadiane oscillator, reguleert indirect de Hmgcs2-expressie (Chavan et al., 2016). Recente waarnemingen geven aan dat extrahepatische tumor-geïnduceerde interleukine-6 ketogenese via PPAR? onderdrukking (Flint et al., 2016).
HMGCS2-enzymactiviteit wordt gereguleerd door meerdere PTM's. HMGCS2-serinefosforylering verhoogde de activiteit ervan in vitro (Grimsrud et al., 2012). HMGCS2-activiteit wordt allosterisch geremd door succinyl-CoA en lysineresidusuccinylatie (Arias et al., 1995; Hegardt, 1999; Lowe en Tubbs, 1985; Quant et al., 1990; Rardin et al., 2013; Reed et al., 1975; Thumelin et al., 1993). Succinylering van HMGCS2-, HMGCL- en BDH1-lysineresiduen in levermitochondriën zijn doelwitten van het NAD+-afhankelijke deacylase sirtuin 5 (SIRT5) (Rardin et al., 2013). HMGCS2-activiteit wordt ook versterkt door SIRT3-lysinedeacetylering, en het is mogelijk dat overspraak tussen acetylering en succinylering de HMGCS2-activiteit reguleert (Rardin et al., 2013; Shimazu et al., 2013). Ondanks het vermogen van deze PTM's om HMGCS2 Km en Vmax te reguleren, zijn fluctuaties van deze PTM's nog niet zorgvuldig in kaart gebracht en zijn ze niet bevestigd als mechanistische aanjagers van ketogenese in vivo.
SCOT komt tot expressie in alle zoogdiercellen die mitochondriën herbergen, behalve die van hepatocyten. Het belang van SCOT-activiteit en ketolyse werd aangetoond in SCOT-KO-muizen, die binnen 48 uur na de geboorte uniforme letaliteit vertoonden als gevolg van hyperketonemische hypoglykemie (Cotter et al., 2011). Weefselspecifiek verlies van SCOT in neuronen of skeletmyocyten veroorzaakt metabole afwijkingen tijdens uithongering, maar is niet dodelijk (Cotter et al., 2013b). Bij mensen manifesteert SCOT-deficiëntie zich vroeg in het leven met ernstige ketoacidose, die lethargie, braken en coma veroorzaakt (Berry et al., 2001; Fukao et al., 2000; Kassovska-Bratinova et al., 1996; Niezen-Koning et al. , 1997; Saudubray et al., 1987; Snyderman et al., 1998; Tildon en Cornblath, 1972). Op cellulair niveau is relatief weinig bekend over SCOT-gen- en eiwitexpressieregulatoren. Oxct1-mRNA-expressie en SCOT-eiwit en -activiteit zijn verminderd in ketotische toestanden, mogelijk door PPAR-afhankelijke mechanismen (Fenselau en Wallis, 1974; Fenselau en Wallis, 1976; Grinblat et al., 1986; Okuda et al., 1991; Turko et al. ., 2001; Wentz et al., 2010). Bij diabetische ketoacidose wordt de mismatch tussen hepatische ketogenese en extrahepatische oxidatie verergerd door een verslechtering van de SCOT-activiteit. Overexpressie van insuline-onafhankelijke glucosetransporter (GLUT1/SLC2A1) in cardiomyocyten remt ook de Oxct1-genexpressie en reguleert de terminale oxidatie van ketonen in een niet-ketotische toestand (Yan et al., 2009). In de lever wordt de overvloed aan Oxct1-mRNA onderdrukt door microRNA-122 en histonmethylering H3K27me3 die duidelijk zijn tijdens de overgang van de foetale naar de neonatale periode (Thorrez et al., 2011). De onderdrukking van Oxct1-expressie in de postnatale periode is echter voornamelijk toe te schrijven aan de evacuatie van hematopoëtische voorlopers die Oxct1 tot expressie brengen uit de lever, in plaats van een verlies van eerder bestaande Oxct1-expressie in terminaal gedifferentieerde hepatocyten. In feite is de expressie van Oxct1-mRNA en SCOT-eiwit in gedifferentieerde hepatocyten extreem laag (Orii et al., 2008).
SCOT wordt ook gereguleerd door PTM's. Het enzym is hypergeacetyleerd in de hersenen van SIRT3 KO-muizen, die ook een verminderde AcAc-afhankelijke acetyl-CoA-productie vertonen (Dittenhafer-Reed et al., 2015). Niet-enzymatische nitrering van tyrosineresiduen van SCOT verzwakt ook de activiteit ervan, wat is gemeld in de harten van verschillende diabetische muizenmodellen (Marcondes et al., 2001; Turko et al., 2001; Wang et al., 2010a). Daarentegen verhoogt nitratie van tryptofaanresidu de SCOT-activiteit (Br�g�re et al., 2010; Rebrin et al., 2007). Moleculaire mechanismen van residu-specifieke nitrering of de-nitratie die zijn ontworpen om SCOT-activiteit te moduleren, kunnen bestaan en moeten worden opgehelderd.
Controverses in extrahepatische ketogenese
Bij zoogdieren is het primaire ketogene orgaan de lever, en alleen hepatocyten en darmepitheelcellen brengen de mitochondriale isovorm van HMGCS2 overvloedig tot expressie (Cotter et al., 2013a; Cotter et al., 2014; McGarry en Foster, 1980; Robinson en Williamson, 1980) . Anaërobe bacteriële fermentatie van complexe polysachariden levert butyraat op, dat wordt geabsorbeerd door colonocyten bij zoogdieren voor terminale oxidatie of ketogenese (Cherbuy et al., 1995), wat een rol kan spelen bij colonocytdifferentiatie (Wang et al., 2016). Met uitzondering van darmepitheelcellen en hepatocyten, is HMGCS2 bijna afwezig in bijna alle andere zoogdiercellen, maar het vooruitzicht van extrahepatische ketogenese is gerezen in tumorcellen, astrocyten van het centrale zenuwstelsel, de nier, pancreas? cellen, retinaal pigmentepitheel (RPE) en zelfs in skeletspieren (Adijanto et al., 2014; Avogaro et al., 1992; El Azzouny et al., 2016; Grabacka et al., 2016; Kang et al., 2015 ; Le Foll et al., 2014; Nonaka et al., 2016; Takagi et al., 2016a; Thevenet et al., 2016; Zhang et al., 2011). Ectopische HMGCS2 is waargenomen in weefsels die geen netto ketogeen vermogen hebben (Cook et al., 2016; Wentz et al., 2010), en HMGCS2 vertoont prospectieve ketogenese-onafhankelijke "moonlighting"-activiteiten, ook in de celkern (Chen et al. , 2016; Kostiuk et al., 2010; Meertens et al., 1998).
Elk extrahepatisch weefsel dat ketonlichamen oxideert, heeft ook het potentieel om ketonlichamen te accumuleren via HMGCS2-onafhankelijke mechanismen (Fig. 2A). Er is echter geen extrahepatisch weefsel waarin een stabiele ketonlichaamconcentratie hoger is dan die in de bloedsomloop (Cotter et al., 2011; Cotter et al., 2013b; Harrison en Long, 1940), wat erop wijst dat ketonlichamen naar beneden worden getransporteerd. concentratiegradiënt via MCT1/2-afhankelijke mechanismen. Een mechanisme van schijnbare extrahepatische ketogenese kan in feite de relatieve verslechtering van ketonoxidatie weerspiegelen. Aanvullende mogelijke verklaringen vallen binnen het domein van de vorming van ketonlichamen. Ten eerste kan de novo ketogenese plaatsvinden via omkeerbare enzymatische activiteit van thiolase en SCOT (Weidemann en Krebs, 1969). Wanneer de concentratie van acetyl-CoA relatief hoog is, verlopen de reacties die normaal verantwoordelijk zijn voor AcAc-oxidatie in de omgekeerde richting (GOLDMAN, 1954). Een tweede mechanisme treedt op wanneer van ?-oxidatie afgeleide tussenproducten accumuleren als gevolg van een TCA-cyclusknelpunt, AcAc-CoA wordt omgezet in l-?OHB-CoA via een reactie die wordt gekatalyseerd door mitochondriaal 3-hydroxyacyl-CoA-dehydrogenase, en verder door 3-hydroxybutyryl CoA-deacylase tot l-?OHB, dat door massaspectrometrie of resonantiespectroscopie niet te onderscheiden is van het fysiologische enantiomeer d-?OHB (Reed en Ozand, 1980). l-?OHB kan chromatografisch of enzymatisch worden onderscheiden van d-?OHB en is aanwezig in extrahepatische weefsels, maar niet in lever of bloed (Hsu et al., 2011). Hepatische ketogenese produceert alleen d-?OHB, het enige enantiomeer dat een BDH-substraat is (Ito et al., 1984; Lincoln et al., 1987; Reed en Ozand, 1980; Scofield et al., 1982; Scofield et al., 1982). Een derde HMGCS2-onafhankelijk mechanisme genereert d-?OHB via aminozuurkatabolisme, met name dat van leucine en lysine. Een vierde mechanisme is alleen duidelijk omdat het te wijten is aan een etiketteringsartefact en daarom pseudoketogenese wordt genoemd. Dit fenomeen is toe te schrijven aan de omkeerbaarheid van de SCOT- en thiolasereacties, en kan een overschatting van de ketonlichaamturnover veroorzaken als gevolg van de isotopische verdunning van ketonlichaamtracer in extrahepatisch weefsel (Des Rosiers et al., 1990; Fink et al., 1988) . Niettemin kan pseudoketogenese in de meeste contexten verwaarloosbaar zijn (Bailey et al., 1990; Keller et al., 1978). Een schema (Fig. 2A) geeft een bruikbare benadering aan om toe te passen, rekening houdend met verhoogde weefsel-steady-state-concentratie van ketonen.
Nier heeft onlangs aandacht gekregen als een potentieel ketogeen orgaan. In de overgrote meerderheid van de staten is de nier een netto-verbruiker van uit de lever afgeleide ketonlichamen, die ketonlichamen uit de bloedbaan uitscheiden of opnieuw opnemen, en de nier is over het algemeen geen netto-ketonlichaamgenerator of -concentrator (Robinson en Williamson, 1980). De auteurs van een klassieke studie concludeerden dat minimale renale ketogenese, gekwantificeerd in een kunstmatig experimenteel systeem, fysiologisch niet relevant was (Weidemann en Krebs, 1969). Onlangs is renale ketogenese afgeleid in diabetische en autofagie-deficiënte muismodellen, maar het is waarschijnlijker dat verschuivingen van meerdere organen in metabole homeostase het integratieve ketonmetabolisme veranderen door input op meerdere organen (Takagi et al., 2016a; Takagi et al., 2016b; Zhang et al., 2011). Een recente publicatie suggereerde renale ketogenese als een beschermend mechanisme tegen ischemie-reperfusieschade in de nier (Tran et al., 2016). Absolute steady-state-concentraties van ?OHB uit extracten van nierweefsel van muizen werden gemeld bij ~4-12 mM. Om te testen of dit houdbaar was, hebben we ?OHB-concentraties gekwantificeerd in nierextracten van gevoede en 24 uur gevaste muizen. Serum ?OHB-concentraties stegen van ~ 100 �M tot 2 mM met 24 uur vasten (Fig. 2B), terwijl renale steady-state ?OHB-concentraties 100 M benaderen in gevoede toestand, en slechts 1 mM in 24 uur vasten (Fig. 2C�E), waarnemingen die consistent zijn met concentraties die meer dan 45 jaar geleden zijn gekwantificeerd (Hems en Brosnan, 1970). Het blijft mogelijk dat in ketotische toestanden van de lever afgeleide ketonlichamen renoprotectief kunnen zijn, maar het bewijs voor renale ketogenese vereist verdere onderbouwing. Overtuigend bewijs dat echte extrahepatische ketogenese ondersteunt, werd gepresenteerd in RPE (Adijanto et al., 2014). Er werd gesuggereerd dat deze intrigerende metabolische transformatie mogelijk van RPE afgeleide ketonen mogelijk zou maken om naar fotoreceptor- of Müller glia-cellen te stromen, wat zou kunnen helpen bij de regeneratie van het buitenste segment van de fotoreceptor.
?OHB als signaleringsbemiddelaar
Hoewel ze energetisch rijk zijn, oefenen ketonlichamen provocerende "niet-canonieke" signaalrollen uit in cellulaire homeostase (Fig. 3) (Newman en Verdin, 2014; Rojas-Morales et al., 2016). ?OHB remt bijvoorbeeld klasse I HDAC's, wat de histonacetylering verhoogt en daardoor de expressie induceert van genen die oxidatieve stress inperken (Shimazu et al., 2013). ?OHB zelf is een covalente histonmodificator van lysineresiduen in levers van nuchtere of door streptozotocine geïnduceerde diabetische muizen (Xie et al., 2016) (zie ook hieronder, De integratie van ketonlichaammetabolisme, post-translationele modificatie en celfysiologie, en ketonlichamen, oxidatieve stress en neuroprotectie).
ï ¿½
?OHB is ook een effector via G-eiwit gekoppelde receptoren. Door onduidelijke moleculaire mechanismen onderdrukt het de activiteit van het sympathische zenuwstelsel en vermindert het het totale energieverbruik en de hartslag door de korteketenvetzuursignalering via G-eiwitgekoppelde receptor 41 (GPR41) te remmen (Kimura et al., 2011). Een van de meest bestudeerde signaaleffecten van ?OHB verloopt via GPR109A (ook bekend als HCAR2), een lid van de hydrocarbonzuur GPCR-subfamilie die tot expressie wordt gebracht in vetweefsel (wit en bruin) (Tunaru et al., 2003), en in immuuncellen (Ahmed et al., 2009). ?OHB is het enige bekende endogene ligand van de GPR109A-receptor (EC50 ~770 �M) geactiveerd door d-?OHB, l-?OHB en butyraat, maar niet AcAc (Taggart et al., 2005). De hoge concentratiedrempel voor GPR109A-activering wordt bereikt door het volgen van een ketogeen dieet, uithongering of tijdens ketoacidose, wat leidt tot remming van de lipolyse van vetweefsel. Het anti-lipolytische effect van GPR109A verloopt door remming van adenylylcyclase en verlaagde cAMP, waardoor hormoongevoelige triglyceridelipase wordt geremd (Ahmed et al., 2009; Tunaru et al., 2003). Dit creëert een negatieve feedbacklus waarin ketose een modulerende rem op ketogenese zet door de afgifte van niet-veresterde vetzuren uit adipocyten te verminderen (Ahmed et al., 2009; Taggart et al., 2005), een effect dat kan worden gecompenseerd door de sympathische drive die lipolyse stimuleert. Niacine (vitamine B3, nicotinezuur) is een krachtige (EC50 ~ 0.1 �M) ligand voor GRP109A, die al tientallen jaren effectief wordt gebruikt voor dyslipidemieën (Benyo et al., 2005; Benyo et al., 2006; Fabbrini et al., 2010a; Lukasova et al., 2011; Tunaru et al., 2003). Terwijl niacine het omgekeerde cholesteroltransport in macrofagen verbetert en atherosclerotische laesies vermindert (Lukasova et al., 2011), blijven de effecten van ?OHB op atherosclerotische laesies onbekend. Hoewel de GPR109A-receptor een beschermende rol speelt en er intrigerende verbanden bestaan tussen het gebruik van ketogeen dieet bij beroerte en neurodegeneratieve ziekten (Fu et al., 2015; Rahman et al., 2014), is een beschermende rol van ?OHB via GPR109A niet in vivo aangetoond .
Ten slotte kan ?OHB de eetlust en verzadiging beïnvloeden. Een meta-analyse van onderzoeken die de effecten van ketogene en zeer energiezuinige diëten hebben gemeten, concludeerde dat deelnemers die deze diëten consumeerden een hogere verzadiging vertoonden in vergelijking met controlediëten (Gibson et al., 2015). Een plausibele verklaring voor dit effect zijn echter de extra metabole of hormonale elementen die de eetlust kunnen moduleren. Muizen die op een ketogeen dieet van knaagdieren werden gehouden, vertoonden bijvoorbeeld een verhoogd energieverbruik in vergelijking met muizen die met chow-controle werden gevoed, ondanks een vergelijkbare calorie-inname, en circulerend leptine of genen van peptiden die het voedingsgedrag reguleren, waren niet veranderd (Kennedy et al., 2007). Een van de voorgestelde mechanismen die de onderdrukking van eetlust door ?OHB suggereren, omvat zowel signalering als oxidatie (Laeger et al., 2010). Hepatocyt-specifieke deletie van het circadiane ritme-gen (Per2) en chromatine-immunoprecipitatie-onderzoeken onthulden dat PER2 het Cpt1a-gen direct activeert en indirect Hmgcs2 reguleert, wat leidt tot verminderde ketose bij Per2-knock-outmuizen (Chavan et al., 2016). Deze muizen vertoonden een verminderde anticipatie op voedsel, die gedeeltelijk werd hersteld door systemische toediening van ?OHB. Toekomstige studies zullen nodig zijn om te bevestigen dat het centrale zenuwstelsel een direct ?OHB-doelwit is, en of ketonoxidatie vereist is voor de waargenomen effecten, of dat er een ander signaalmechanisme bij betrokken is. Andere onderzoekers hebben de mogelijkheid ingeroepen van lokale door astrocyten afgeleide ketogenese in de ventromediale hypothalamus als een regulator van de voedselinname, maar deze voorlopige waarnemingen zullen ook profiteren van genetische en op flux gebaseerde beoordelingen (Le Foll et al., 2014). De relatie tussen ketose en tekort aan voedingsstoffen blijft van belang omdat honger en verzadiging belangrijke elementen zijn bij mislukte pogingen om gewicht te verliezen.
Integratie van ketonlichaammetabolisme, post-translationele modificatie en celfysiologie
Ketonlichamen dragen bij aan gecompartimentaliseerde pools van acetyl-CoA, een belangrijk tussenproduct dat een prominente rol speelt in het cellulaire metabolisme (Pietrocola et al., 2015). Een rol van acetyl-CoA is om te dienen als een substraat voor acetylering, een enzymatisch gekatalyseerde covalente histonmodificatie (Choudhary et al., 2014; Dutta et al., 2016; Fan et al., 2015; Menzies et al., 2016 ). Een groot aantal dynamisch geacetyleerde mitochondriale eiwitten, waarvan er vele kunnen voorkomen via niet-enzymatische mechanismen, zijn ook voortgekomen uit computationele proteomics-onderzoeken (Dittenhafer-Reed et al., 2015; Hebert et al., 2013; Rardin et al., 2013 ; Shimazu et al., 2010). Lysine-deacetylasen gebruiken een zink-cofactor (bijv. nucleocytosolic HDAC's) of NAD+ als co-substraat (sirtuins, SIRT's) (Choudhary et al., 2014; Menzies et al., 2016). Het acetylproteoom dient als zowel sensor als effector van de totale cellulaire acetyl-CoA-pool, aangezien fysiologische en genetische manipulaties elk resulteren in niet-enzymatische globale variaties van acetylering (Weinert et al., 2014). Aangezien intracellulaire metabolieten dienen als modulatoren van acetylering van lysineresiduen, is het belangrijk om de rol van ketonlichamen te overwegen, waarvan de overvloed zeer dynamisch is.
?OHB is een epigenetische modifier via ten minste twee mechanismen. Verhoogde ?OHB-spiegels veroorzaakt door vasten, caloriebeperking, directe toediening of langdurige inspanning veroorzaken HDAC-remming of histonacetyltransferase-activering (Marosi et al., 2016; Sleiman et al., 2016) of tot oxidatieve stress (Shimazu et al., 2013) . ?OHB-remming van HDAC3 zou de metabole fysiologie van pasgeborenen kunnen reguleren (Rando et al., 2016). Onafhankelijk modificeert ?OHB zelf histonlysineresiduen direct (Xie et al., 2016). Langdurig vasten of door steptozotocine geïnduceerde diabetische ketoacidose verhoogde histon-?-hydroxybutyrylering. Hoewel het aantal lysine-a-hydroxybutyrylerings- en acetyleringsplaatsen vergelijkbaar was, werd stoichiometrisch grotere histon-a-hydroxybutyrylering dan acetylering waargenomen. Verschillende genen werden beïnvloed door histonlysine ?-hydroxybutyrylering, versus acetylering of methylering, wat verschillende cellulaire functies suggereert. Of ?-hydroxybutyrylering spontaan of enzymatisch is, is niet bekend, maar breidt het bereik van mechanismen uit via ketonlichamen die transcriptie dynamisch beïnvloeden.
Essentiële celherprogrammering tijdens caloriebeperking en nutriëntdeprivatie kan worden gemedieerd in respectievelijk SIRT3- en SIRT5-afhankelijke mitochondriale deacetylering en desuccinylering, die ketogene en ketolytische eiwitten reguleren op post-translationeel niveau in lever- en extrahepatische weefsels (Dittenhafer-Reed et al., 2015; Hebert et al., 2013; Rardin et al., 2013; Shimazu et al., 2010). Hoewel stoichiometrische vergelijking van bezette locaties niet noodzakelijk direct verband houdt met verschuivingen in metabole flux, is mitochondriale acetylering dynamisch en kan deze worden aangedreven door acetyl-CoA-concentratie of mitochondriale pH, in plaats van enzymatische acetyltransferasen (Wagner en Payne, 2013). Dat SIRT3 en SIRT5 de activiteiten van enzymen die het ketonlichaam metaboliseren moduleren, roept de vraag op naar de wederkerige rol van ketonen bij het vormen van het acetylproteoom, succinylproteoom en andere dynamische cellulaire doelen. Aangezien variaties van ketogenese NAD+-concentraties weerspiegelen, zouden de ketonproductie en overvloed de sirtuin-activiteit kunnen reguleren, waardoor de totale acetyl-CoA / succinyl-CoA-pools, het acylproteoom en dus de mitochondriale en celfysiologie worden beïnvloed. ?-hydroxybutyrylering van enzymlysineresiduen zou een nieuwe laag kunnen toevoegen aan cellulaire herprogrammering. In extrahepatische weefsels kan ketonlichaamoxidatie analoge veranderingen in celhomeostase stimuleren. Hoewel de compartimentering van acetyl-CoA-pools sterk gereguleerd is en een breed spectrum van cellulaire veranderingen coördineert, vereist het vermogen van ketonlichamen om zowel mitochondriale als cytoplasmatische acetyl-CoA-concentraties direct vorm te geven opheldering (Chen et al., 2012; Corbet et al., 2016; Pougovkina et al., 2014; Schwer et al., 2009; Wellen en Thompson, 2012). Omdat acetyl-CoA-concentraties strak gereguleerd zijn en acetyl-CoA membraan-impermeabel is, is het van cruciaal belang om rekening te houden met de drijvende mechanismen die de homeostase van acetyl-CoA coördineren, inclusief de productiesnelheden en terminale oxidatie in de TCA-cyclus, omzetting in ketonlichamen, mitochondriale efflux via carnitine-acetyltransferase (CrAT), of acetyl-CoA-export naar cytosol na conversie naar citraat en afgifte door ATP-citraatlyase (ACLY). De sleutelrollen van deze laatste mechanismen in celacetylproteoom en homeostase vereisen een op elkaar afgestemd begrip van de rollen van ketogenese en ketonoxidatie (Das et al., 2015; McDonnell et al., 2016; Moussaieff et al., 2015; Overmyer et al., 2015; Seiler et al., 2014; Seiler et al., 2015; Wellen et al., 2009; Wellen en Thompson, 2012). Convergente technologieën in metabolomics en acylproteomics in de setting van genetisch gemanipuleerde modellen zullen nodig zijn om doelen en resultaten te specificeren.
Anti- en pro-inflammatoire reacties op ketonlichamen
Ketose en ketonlichamen moduleren ontsteking en immuuncelfunctie, maar er zijn verschillende en zelfs tegenstrijdige mechanismen voorgesteld. Langdurig gebrek aan voedingsstoffen vermindert ontstekingen (Youm et al., 2015), maar de chronische ketose van diabetes type 1 is een pro-inflammatoire toestand (Jain et al., 2002; Kanikarla-Marie en Jain, 2015; Kurepa et al., 2012 ). Op mechanisme gebaseerde signaleringsrollen voor ?OHB bij ontstekingen komen naar voren omdat veel cellen van het immuunsysteem, waaronder macrofagen of monocyten, GPR109A overvloedig tot expressie brengen. Hoewel ?OHB een overwegend ontstekingsremmende reactie uitoefent (Fu et al., 2014; Gambhir et al., 2012; Rahman et al., 2014; Youm et al., 2015), kunnen hoge concentraties ketonlichamen, met name AcAc, mogelijk een pro-inflammatoire respons veroorzaken (Jain et al., 2002; Kanikarla-Marie en Jain, 2015; Kurepa et al., 2012).
Ontstekingsremmende rollen van GPR109A-liganden bij atherosclerose, obesitas, inflammatoire darmaandoeningen, neurologische aandoeningen en kanker zijn beoordeeld (Graff et al., 2016). GPR109A-expressie wordt verhoogd in RPE-cellen van diabetische modellen, menselijke diabetespatiënten (Gambhir et al., 2012) en in microglia tijdens neurodegeneratie (Fu et al., 2014). Ontstekingsremmende effecten van ?OHB worden versterkt door overexpressie van GPR109A in RPE-cellen en opgeheven door farmacologische remming of genetische knock-out van GPR109A (Gambhir et al., 2012). ?OHB en exogeen nicotinezuur (Taggart et al., 2005), beide verlenen ontstekingsremmende effecten in TNF? of LPS-geïnduceerde ontsteking door het verlagen van de niveaus van pro-inflammatoire eiwitten (iNOS, COX-2) of uitgescheiden cytokines (TNF?, IL-1?, IL-6, CCL2/MCP-1), gedeeltelijk door remming van NF -?B-translocatie (Fu et al., 2014; Gambhir et al., 2012). ?OHB vermindert ER-stress en het NLRP3-inflammasoom, waardoor de antioxidatieve stressrespons wordt geactiveerd (Bae et al., 2016; Youm et al., 2015). Bij neurodegeneratieve ontsteking omvat GPR109A-afhankelijke ?OHB-gemedieerde bescherming echter geen ontstekingsmediatoren zoals MAPK-routesignalering (bijv. ERK, JNK, p38) (Fu et al., 2014), maar vereist mogelijk COX-1-afhankelijke PGD2 productie (Rahman et al., 2014). Het is intrigerend dat macrofaag GPR109A nodig is om een neuroprotectief effect uit te oefenen in een ischemisch beroertemodel (Rahman et al., 2014), maar het vermogen van ?OHB om het NLRP3-inflammasoom in uit beenmerg afgeleide macrofagen te remmen is GPR109A-onafhankelijk (Youm et al., 2015). ., 2014). Hoewel de meeste onderzoeken ?OHB koppelen aan ontstekingsremmende effecten, kan ?OHB pro-inflammatoir zijn en markers van lipideperoxidatie in kalfhepatocyten verhogen (Shi et al., XNUMX). Anti- versus pro-inflammatoire effecten van ?OHB kunnen dus afhangen van celtype, ?OHB-concentratie, blootstellingsduur en de aan- of afwezigheid van co-modulatoren.
In tegenstelling tot ?OHB kan AcAc pro-inflammatoire signalering activeren. Verhoogd AcAc, vooral met een hoge glucoseconcentratie, intensiveert endotheelcelbeschadiging door een NADPH-oxidase / oxidatieve stress-afhankelijk mechanisme (Kanikarla-Marie en Jain, 2015). Hoge AcAc-concentraties in de navelstreng van diabetische moeders waren gecorreleerd met een hogere eiwitoxidatiesnelheid en MCP-1-concentratie (Kurepa et al., 2012). Hoge AcAc bij diabetespatiënten was gecorreleerd met TNF? expressie (Jain et al., 2002), en AcAc, maar niet ?OHB, induceerde TNF?, MCP-1-expressie, ROS-accumulatie en verminderd cAMP-niveau in U937 menselijke monocytcellen (Jain et al., 2002; Kurepa et al. ., 2012).
Ketonlichaamafhankelijke signaleringsverschijnselen worden vaak alleen geactiveerd bij hoge ketonlichaamconcentraties (> 5 mM), en in het geval van veel onderzoeken die ketonen koppelen aan pro- of ontstekingsremmende effecten, door onduidelijke mechanismen. Vanwege de tegenstrijdige effecten van ?OHB versus AcAc op ontstekingen en het vermogen van de AcAc/?OHB-verhouding om het mitochondriale redoxpotentieel te beïnvloeden, vergelijken de beste experimenten die de rol van ketonlichamen op cellulaire fenotypen beoordelen bovendien de effecten van AcAc en ? OHB in verschillende verhoudingen en in verschillende cumulatieve concentraties [bijv. (Saito et al., 2016)]. Ten slotte kan AcAc alleen in de handel worden gekocht als een lithiumzout of als een ethylester die vóór gebruik basehydrolyse vereist. Lithiumkation induceert onafhankelijk signaaltransductiecascades (Manji et al., 1995), en AcAc-anion is labiel. Ten slotte kunnen studies met racemisch d/l-?OHB verwarrend zijn, aangezien alleen het d-?OHB-stereo-isomeer kan worden geoxideerd tot AcAc, maar d-?OHB en l-?OHB kunnen elk signaal via GPR109A, het NLRP3-inflammasoom remmen, en dienen als lipogene substraten.
Ketonlichamen, oxidatieve stress en neuroprotectie
Oxidatieve stress wordt typisch gedefinieerd als een toestand waarin ROS in overmaat wordt gepresenteerd, als gevolg van overmatige productie en/of verminderde eliminatie. Antioxidant en oxidatieve stressverlagende rollen van ketonlichamen zijn zowel in vitro als in vivo uitgebreid beschreven, met name in de context van neuroprotectie. Aangezien de meeste neuronen niet effectief hoogenergetische fosfaten uit vetzuren genereren, maar ketonlichamen oxideren wanneer er een tekort aan koolhydraten is, zijn neuroprotectieve effecten van ketonlichamen bijzonder belangrijk (Cahill GF Jr, 2006; Edmond et al., 1987; Yang et al., 1987). In modellen voor oxidatieve stress suggereren BDH1-inductie en SCOT-onderdrukking dat het metabolisme van het ketonlichaam opnieuw kan worden geprogrammeerd om verschillende celsignalering, redoxpotentiaal of metabolische vereisten te ondersteunen (Nagao et al., 2016; Tieu et al., 2003).
Ketonlichamen verminderen de gradaties van cellulaire schade, verwonding, dood en lagere apoptose in neuronen en cardiomyocyten (Haces et al., 2008; Maalouf et al., 2007; Nagao et al., 2016; Tieu et al., 2003). Ingeroepen mechanismen zijn gevarieerd en niet altijd lineair gerelateerd aan concentratie. Lage millimolaire concentraties van (d of l)-?OHB vangen ROS (hydroxylanion), terwijl AcAc talrijke ROS-soorten opruimt, maar alleen bij concentraties die het fysiologische bereik overschrijden (IC50 20-67 mM) (Haces et al., 2008) . Omgekeerd is een gunstige invloed op de redoxpotentiaal van de elektronentransportketen een mechanisme dat gewoonlijk wordt gekoppeld aan d-?OHB. Terwijl alle drie de ketonlichamen (d/l-?OHB en AcAc) neuronale celdood en ROS-accumulatie veroorzaakten door chemische remming van glycolyse verminderden, voorkwamen alleen d-?OHB en AcAc neuronale ATP-afname. Omgekeerd, in een hypoglycemisch in vivo-model, verhinderde (d of l)-?OHB, maar niet AcAc, hippocampale lipideperoxidatie (Haces et al., 2008; Maalouf et al., 2007; Marosi et al., 2016; Murphy, 2009 ; Tieu et al., 2003). In vivo studies van muizen die een ketogeen dieet kregen (87% kcal vet en 13% eiwit) vertoonden neuroanatomische variatie in antioxidantcapaciteit (Ziegler et al., 2003), waarbij de meest ingrijpende veranderingen werden waargenomen in de hippocampus, met een toename van glutathionperoxidase en totaal antioxidant capaciteiten.
Ketogeen dieet, ketonesters (zie ook Therapeutisch gebruik van ketogeen dieet en exogene ketonlichamen), of toediening van ?OHB oefenen neuroprotectie uit in modellen van ischemische beroerte (Rahman et al., 2014); de ziekte van Parkinson (Tieu et al., 2003); zuurstofvergiftiging van het centrale zenuwstelsel (D'Agostino et al., 2013); epileptische spasmen (Yum et al., 2015); mitochondriale encefalomyopathie, lactaatacidose en beroerte-achtige (MELAS) episodes-syndroom (Frey et al., 2016) en de ziekte van Alzheimer (Cunnane en Crawford, 2003; Yin et al., 2016). Omgekeerd toonde een recent rapport histopathologisch bewijs aan van neurodegeneratieve progressie door een ketogeen dieet in een transgeen muismodel van abnormaal mitochondriaal DNA-herstel, ondanks toename van mitochondriale biogenese en antioxidantsignaturen (Lauritzen et al., 2016). Andere tegenstrijdige rapporten suggereren dat blootstelling aan hoge ketonlichaamconcentraties oxidatieve stress veroorzaakt. Hoge ?OHB- of AcAc-doses induceerden stikstofmonoxide-secretie, lipideperoxidatie, verminderde expressie van SOD, glutathionperoxidase en katalase in hepatocyten van kalveren, terwijl in hepatocyten van ratten de MAPK-pathway-inductie werd toegeschreven aan AcAc maar niet aan ?OHB (Abdelmegeed et al., 2004 ; Shi et al., 2014; Shi et al., 2016).
Alles bij elkaar genomen, koppelen de meeste rapporten ?OHB aan de vermindering van oxidatieve stress, aangezien de toediening ervan de productie van ROS/superoxide remt, lipideperoxidatie en eiwitoxidatie voorkomt, de antioxidant-eiwitniveaus verhoogt en de mitochondriale ademhaling en ATP-productie verbetert (Abdelmegeed et al., 2004; Haces et al., 2008; Jain et al., 1998; Jain et al., 2002; Kanikarla-Marie en Jain, 2015; Maalouf et al., 2007; Maalouf en Rho, 2008; Marosi et al., 2016; Tieu et al., 2003; Yin et al., 2016; Ziegler et al., 2003). Hoewel AcAc directer gecorreleerd is dan ?OHB met de inductie van oxidatieve stress, zijn deze effecten niet altijd gemakkelijk te onderscheiden van toekomstige pro-inflammatoire reacties (Jain et al., 2002; Kanikarla-Marie en Jain, 2015; Kanikarla-Marie en Jaïn, 2016). Bovendien is het van cruciaal belang om te bedenken dat het schijnbare antioxidatieve voordeel dat wordt verleend door pleiotrope ketogene diëten mogelijk niet wordt getransduceerd door ketonlichamen zelf, en dat neuroprotectie die wordt verleend door ketonlichamen mogelijk niet volledig te wijten is aan oxidatieve stress. Bijvoorbeeld tijdens glucosedeprivatie, in een model van glucosedeprivatie in corticale neuronen, stimuleerde ?OHB autofagische flux en verhinderde autofagosoomaccumulatie, wat geassocieerd was met verminderde neuronale dood (Camberos-Luna et al., 2016). d-?OHB induceert ook de canonieke antioxidant-eiwitten FOXO3a, SOD, MnSOD en catalase, prospectief door HDAC-remming (Nagao et al., 2016; Shimazu et al., 2013).
Niet-alcoholische leververvetting (NAFLD) en ketonenmetabolisme
Obesitas-geassocieerde NAFLD en niet-alcoholische steatohepatitis (NASH) zijn de meest voorkomende oorzaken van leverziekte in westerse landen (Rinella en Sanyal, 2016), en NASH-geïnduceerd leverfalen is een van de meest voorkomende redenen voor levertransplantatie. Hoewel overmatige opslag van triacylglycerolen in hepatocyten >5% van het levergewicht (NAFL) alleen geen degeneratieve leverfunctie veroorzaakt, correleert de progressie naar NAFLD bij mensen met systemische insulineresistentie en een verhoogd risico op type 2-diabetes, en kan het bijdragen aan de pathogenese van hart- en vaatziekten en chronische nierziekte (Fabbrini et al., 2009; Targher et al., 2010; Targher en Byrne, 2013). De pathogene mechanismen van NAFLD en NASH zijn niet volledig begrepen, maar omvatten afwijkingen van het hepatocytmetabolisme, hepatocyt-autofagie en endoplasmatisch reticulumstress, leverimmuuncelfunctie, vetweefselontsteking en systemische inflammatoire mediatoren (Fabbrini et al., 2009; Masuoka en Chalasani, 2013 ; Targher et al., 2010; Yang et al., 2010). Verstoringen van het koolhydraat-, lipide- en aminozuurmetabolisme komen voor in en dragen bij aan obesitas, diabetes en NAFLD bij mensen en in modelorganismen [besproken in (Farese et al., 2012; Lin en Accili, 2011; Newgard, 2012; Samuel en Shulman, 2012; Sun en Lazar, 2013)]. Hoewel hepatocytafwijkingen in het cytoplasmatisch lipidenmetabolisme vaak worden waargenomen in NAFLD (Fabbrini et al., 2010b), is de rol van het mitochondriale metabolisme, dat de oxidatieve verwijdering van vetten regelt, minder duidelijk in de pathogenese van NAFLD. Afwijkingen van het mitochondriaal metabolisme komen voor in en dragen bij aan de pathogenese van NAFLD/NASH (Hyotylainen et al., 2016; Serviddio et al., 2011; Serviddio et al., 2008; Wei et al., 2008). Er is algemeen (Felig et al., 1974; Iozzo et al., 2010; Koliaki et al., 2015; Satapati et al., 2015; Satapati et al., 2012; Sunny et al., 2011) maar niet uniform ( Koliaki en Roden, 2013; Perry et al., 2016; Rector et al., 2010) consensus dat, voorafgaand aan de ontwikkeling van bonafide NASH, mitochondriale oxidatie in de lever, en in het bijzonder vetoxidatie, wordt vergroot in obesitas, systemische insulineresistentie en NAFLD. Het is waarschijnlijk dat naarmate NAFLD vordert, oxidatieve capaciteitsheterogeniteit, zelfs tussen individuele mitochondriën, naar voren komt, en uiteindelijk de oxidatieve functie wordt aangetast (Koliaki et al., 2015; Rector et al., 2010; Satapati et al., 2008; Satapati et al., 2012; Satapati et al. ., XNUMX).
Ketogenese wordt vaak gebruikt als een proxy voor hepatische vetoxidatie. Verslechteringen van ketogenese komen naar voren naarmate NAFLD vordert in diermodellen, en waarschijnlijk bij mensen. Door onvolledig gedefinieerde mechanismen onderdrukt hyperinsulinemie ketogenese, wat mogelijk bijdraagt aan hypoketonemie in vergelijking met magere controles (Bergman et al., 2007; Bickerton et al., 2008; Satapati et al., 2012; Soeters et al., 2009; Sunny et al. , 2011; Vice et al., 2005). Desalniettemin is het vermogen van circulerende ketonlichaamconcentraties om NAFLD te voorspellen controversieel (Mnnist et al., 2015; Sanyal et al., 2001). Robuuste kwantitatieve magnetische resonantie spectroscopische methoden in diermodellen onthulden een verhoogde keton-turnoversnelheid met matige insulineresistentie, maar verminderde percentages waren duidelijk met ernstigere insulineresistentie (Satapati et al., 2012; Sunny et al., 2010). Bij zwaarlijvige mensen met leververvetting is de ketogene snelheid normaal (Bickerton et al., 2008; Sunny et al., 2011), en daarom neemt de snelheid van ketogenese af ten opzichte van de verhoogde vetzuurbelasting in hepatocyten. Bijgevolg kan van ?-oxidatie afgeleid acetyl-CoA worden gericht op terminale oxidatie in de TCA-cyclus, waardoor terminale oxidatie, door fosfoenolpyruvaat aangedreven gluconeogenese via anaplerose/cataplerose en oxidatieve stress wordt verhoogd. Acetyl-CoA wordt mogelijk ook geëxporteerd vanuit mitochondriën als citraat, een voorlopersubstraat voor lipogenese (Fig. 4) (Satapati et al., 2015; Satapati et al., 2012; Solinas et al., 2015). Terwijl ketogenese minder snel reageert op insuline of vasten bij langdurige obesitas (Satapati et al., 2012), blijven de onderliggende mechanismen en stroomafwaartse gevolgen hiervan onvolledig begrepen. Recent bewijs geeft aan dat mTORC1 ketogenese onderdrukt op een manier die mogelijk stroomafwaarts is van insulinesignalering (Kucejova et al., 2016), wat in overeenstemming is met de waarnemingen dat mTORC1 PPAR?-gemedieerde Hmgcs2-inductie remt (Sengupta et al., 2010) ( zie ook Regeling van HMGCS2 en SCOT/OXCT1).
ï ¿½
Voorlopige observaties van onze groep suggereren nadelige gevolgen voor de lever van ketogene insufficiëntie (Cotter et al., 2014). Om de hypothese te testen dat gestoorde ketogenese, zelfs in koolhydraatrijke en dus 'niet-ketogene' toestanden, bijdraagt aan een abnormaal glucosemetabolisme en steatohepatitis veroorzaakt, hebben we een muismodel met duidelijke ketogene insufficiëntie gegenereerd door toediening van antisense-oligonucleotiden (ASO) gericht op Hmgcs2. Verlies van HMGCS2 in standaard vetarme volwassen muizen veroorzaakte milde hyperglykemie en een duidelijk verhoogde productie van honderden levermetabolieten, waarvan een reeks sterk suggereerde activatie van lipogenese. Vetrijke voeding van muizen met onvoldoende ketogenese resulteerde in uitgebreide hepatocytbeschadiging en -ontsteking. Deze bevindingen ondersteunen de centrale hypothesen dat (i) ketogenese geen passieve overlooproute is, maar eerder een dynamisch knooppunt in hepatische en geïntegreerde fysiologische homeostase, en (ii) voorzichtige ketogene augmentatie om NAFLD/NASH en verstoord leverglucosemetabolisme te onderzoeken, is het waard om te worden onderzocht. .
Hoe kan een verminderde ketogenese bijdragen aan leverbeschadiging en veranderde glucosehomeostase? De eerste overweging is of de boosdoener een tekort aan ketogene flux is, of ketonen zelf. Een recent rapport suggereert dat ketonlichamen door oxidatieve stress veroorzaakte leverbeschadiging kunnen verminderen als reactie op n-3 meervoudig onverzadigde vetzuren (Pawlak et al., 2015). Bedenk dat vanwege een gebrek aan SCOT-expressie in hepatocyten, ketonlichamen niet worden geoxideerd, maar dat ze kunnen bijdragen aan lipogenese en een verscheidenheid aan signaleringsrollen kunnen vervullen, onafhankelijk van hun oxidatie (zie ook niet-oxidatieve metabolische lotgevallen van ketonlichamen en ?OHB als een signaalbemiddelaar). Het is ook mogelijk dat van hepatocyten afgeleide ketonlichamen kunnen dienen als signaal en/of metaboliet voor naburige celtypen in de leveracinus, waaronder stellaatcellen en Kupffer-celmacrofagen. Hoewel de beperkte beschikbare literatuur suggereert dat macrofagen niet in staat zijn ketonlichamen te oxideren, is dit alleen gemeten met behulp van klassieke methodologieën en alleen in peritoneale macrofagen (Newsholme et al., 1986; Newsholme et al., 1987), wat aangeeft dat een re- beoordeling is passend gezien de overvloedige SCOT-expressie in van beenmerg afgeleide macrofagen (Youm et al., 2015).
De ketogene flux van hepatocyten kan ook cytoprotectief zijn. Hoewel heilzame mechanismen mogelijk niet per se afhankelijk zijn van ketogenese, zijn ketogene diëten met weinig koolhydraten in verband gebracht met verbetering van NAFLD (Browning et al., 2011; Foster et al., 2010; Kani et al., 2014; Schugar en Crawford, 2012) . Onze waarnemingen geven aan dat ketogenese van hepatocyten feedback kan geven en de TCA-cyclusflux, anaplerotische flux, van fosfoenolpyruvaat afgeleide gluconeogenese (Cotter et al., 2014) en zelfs glycogeenomzet kan reguleren en reguleren. Ketogene stoornis zorgt ervoor dat acetyl-CoA de TCA-flux verhoogt, wat in de lever is gekoppeld aan verhoogde ROS-gemedieerde schade (Satapati et al., 2015; Satapati et al., 2012); dwingt afleiding van koolstof naar de novo gesynthetiseerde lipidesoorten die cytotoxisch zouden kunnen zijn; en voorkomt heroxidatie van NADH tot NAD+ (Cotter et al., 2014) (Fig. 4). Alles bij elkaar genomen, zijn toekomstige experimenten nodig om mechanismen aan te pakken waardoor relatieve ketogene insufficiëntie onaangepast kan worden, kan bijdragen aan hyperglykemie, steatohepatitis kan veroorzaken, en of deze mechanismen werkzaam zijn in menselijke NAFLD/NASH. Aangezien epidemiologisch bewijs een verminderde ketogenese suggereert tijdens de progressie van steatohepatitis (Embade et al., 2016; Marinou et al., 2011; M�nnist� et al., 2015; Pramfalk et al., 2015; Safaei et al., 2016) therapieën die hepatische ketogenese verhogen, kunnen heilzaam zijn (Degirolamo et al., 2016; Honda et al., 2016).
Ketonlichamen en hartfalen (HF)
Met een stofwisselingssnelheid van meer dan 400 kcal/kg/dag en een omzet van 6 kg ATP/dag, is het hart het orgaan met het hoogste energieverbruik en de meeste oxidatieve vraag (Ashrafian et al., 35; Wang et al., 2007b). De overgrote meerderheid van de energieomzet van het hart bevindt zich in de mitochondriën en 2010% van deze voorraad is afkomstig van FAO. Het hart is omnivoor en flexibel onder normale omstandigheden, maar het pathologisch hermodellerende hart (bijvoorbeeld als gevolg van hypertensie of myocardinfarct) en het diabetische hart worden elk metabool inflexibel (Balasse en Fery, 70; BING, 1989; Fukao et al., 1954 ; Lopaschuk et al., 2004; Taegtmeyer et al., 2010; Taegtmeyer et al., 1980; Young et al., 2002). Inderdaad, genetisch geprogrammeerde afwijkingen van het hartbrandstofmetabolisme in muismodellen veroorzaken cardiomyopathie (Carley et al., 2002; Neubauer, 2014). Onder fysiologische omstandigheden oxideren normale harten ketonlichamen in verhouding tot hun afgifte, ten koste van vetzuur- en glucoseoxidatie, en myocardium is de grootste ketonlichaamverbruiker per massa-eenheid (BING, 2007; Crawford et al., 1954; GARLAND et al. ., 2009; Hasselbaink et al., 1962; Jeffrey et al., 2003; Pelletier et al., 1995; Tardif et al., 2007; Yan et al., 2001). In vergelijking met vetzuuroxidatie zijn ketonlichamen energetisch efficiënter, waardoor er meer energie beschikbaar is voor ATP-synthese per geïnvesteerd zuurstofmolecuul (P/O-verhouding) (Kashiwaya et al., 2009; Sato et al., 2010; Veech, 1995) . Oxidatie van ketonlichamen levert ook potentieel hogere energie op dan FAO, waardoor ubiquinon geoxideerd blijft, wat de redox-spanwijdte in de elektronentransportketen verhoogt en meer energie beschikbaar maakt om ATP te synthetiseren (Sato et al., 2004; Veech, 1995). Oxidatie van ketonlichamen kan ook de ROS-productie verminderen, en dus oxidatieve stress (Veech, 2004).
Voorlopige interventionele en observationele studies wijzen op een mogelijke heilzame rol van ketonlichamen in het hart. In de experimentele context van ischemie/reperfusieschade verleenden ketonlichamen potentiële cardioprotectieve effecten (Al-Zaid et al., 2007; Wang et al., 2008), mogelijk als gevolg van de toename van mitochondriale overvloed in het hart of opregulatie van cruciale oxidatieve fosforylering bemiddelaars (Snorek et al., 2012; Zou et al., 2002). Recente onderzoeken geven aan dat het gebruik van ketonlichaam wordt verhoogd in falende harten van muizen (Aubert et al., 2016) en mensen (Bedi et al., 2016), wat eerdere observaties bij mensen ondersteunt (BING, 1954; Fukao et al., 2000; Janardhan et al., 2011; Longo et al., 2004; Rudolph en Schinz, 1973; Tildon en Cornblath, 1972). De circulerende ketonlichaamconcentraties zijn verhoogd bij patiënten met hartfalen, in directe verhouding tot de vuldruk, waarnemingen waarvan het mechanisme en de betekenis onbekend blijven (Kupari et al., 1995; Lommi et al., 1996; Lommi et al., 1997; Neely et al. ., 1972), maar muizen met selectieve SCOT-deficiëntie in cardiomyocyten vertonen versnelde pathologische ventriculaire hermodellering en ROS-signaturen als reactie op chirurgisch geïnduceerde schade door drukoverbelasting (Schugar et al., 2014).
Recente intrigerende observaties bij diabetestherapie hebben een mogelijk verband aangetoond tussen myocardiaal ketonmetabolisme en pathologische ventriculaire remodellering (Fig. 5). Remming van de renale proximale tubulaire natrium/glucose-cotransporter 2 (SGLT2i) verhoogt de circulerende ketonlichaamconcentraties bij mensen (Ferrannini et al., 2016a; Inagaki et al., 2015) en muizen (Suzuki et al., 2014) via verhoogde hepatische ketogenese (Ferrannini et al., 2014; Ferrannini et al., 2016a; Katz en Leiter, 2015; Mudaliar et al., 2015). Opvallend is dat ten minste één van deze middelen de HF-hospitalisatie verminderde (bijv. zoals onthuld door de EMPA-REG OUTCOME-studie), en de cardiovasculaire mortaliteit verbeterde (Fitchett et al., 2016; Sonesson et al., 2016; Wu et al., 2016a. ; Zinman et al., 2015). Hoewel de drijvende mechanismen achter gunstige HF-resultaten voor gekoppelde SGLT2i actief worden besproken, is het overlevingsvoordeel waarschijnlijk multifactorieel, prospectief inclusief ketose maar ook heilzame effecten op gewicht, bloeddruk, glucose- en urinezuurniveaus, arteriële stijfheid, het sympathische zenuwstelsel, osmotische diurese / verminderd plasmavolume en verhoogde hematocriet (Raz en Cahn, 2016; Vallon en Thomson, 2016). Samengevat blijft het idee dat therapeutisch toenemende ketonemie, hetzij bij HF-patiënten, of bij patiënten met een hoog risico om HF te ontwikkelen, controversieel, maar wordt actief onderzocht in preklinische en klinische onderzoeken (Ferrannini et al., 2016b; Kolwicz et al., 2016; Lopaschuk en Verma, 2016; Mudaliar et al., 2016; Taegtmeyer, 2016).
ï ¿½
Ketonlichamen in de kankerbiologie
Verbindingen tussen ketonlichamen en kanker komen snel op, maar studies in zowel diermodellen als mensen hebben uiteenlopende conclusies opgeleverd. Omdat ketonmetabolisme dynamisch is en reageert op de voedingstoestand, is het verleidelijk om biologische verbindingen met kanker na te streven vanwege het potentieel voor nauwkeurig geleide voedingstherapieën. Kankercellen ondergaan metabolische herprogrammering om snelle celproliferatie en groei te behouden (DeNicola en Cantley, 2015; Pavlova en Thompson, 2016). Het klassieke Warburg-effect in het metabolisme van kankercellen komt voort uit de dominante rol van glycolyse en melkzuurfermentatie om energie over te dragen en te compenseren voor een lagere afhankelijkheid van oxidatieve fosforylering en beperkte mitochondriale ademhaling (De Feyter et al., 2016; Grabacka et al., 2016; Kang et al., 2015; Poff et al., 2014; Shukla et al., 2014). Glucose-koolstof wordt voornamelijk geleid door glycolyse, de pentosefosfaatroute en lipogenese, die samen zorgen voor tussenproducten die nodig zijn voor de uitbreiding van de tumorbiomassa (Grabacka et al., 2016; Shukla et al., 2014; Yoshii et al., 2015). Aanpassing van kankercellen aan glucosetekort vindt plaats door het vermogen om alternatieve brandstofbronnen te benutten, waaronder acetaat, glutamine en aspartaat (Jaworski et al., 2016; Sullivan et al., 2015). Beperkte toegang tot pyruvaat onthult bijvoorbeeld het vermogen van kankercellen om glutamine om te zetten in acetyl-CoA door carboxylatie, waarbij zowel de energetische als de anabole behoeften behouden blijven (Yang et al., 2014). Een interessante aanpassing van kankercellen is het gebruik van acetaat als brandstof (Comerford et al., 2014; Jaworski et al., 2016; Mashimo et al., 2014; Wright en Simone, 2016; Yoshii et al., 2015). Acetaat is ook een substraat voor lipogenese, wat van cruciaal belang is voor de proliferatie van tumorcellen, en de winst van dit lipogene kanaal wordt geassocieerd met een kortere overleving van de patiënt en een grotere tumorlast (Comerford et al., 2014; Mashimo et al., 2014; Yoshii et al. ., 2015).
Niet-kankercellen verplaatsen hun energiebron gemakkelijk van glucose naar ketonlichamen tijdens glucosetekort. Deze plasticiteit kan meer variabel zijn tussen kankerceltypes, maar in vivo geïmplanteerde hersentumoren oxideerden [2,4-13C2]-?OHB in een vergelijkbare mate als omringend hersenweefsel (De Feyter et al., 2016). 'Reverse Warburg-effect' of 'tumormetabolisme met twee compartimenten' veronderstellen dat kankercellen 'OHB-productie induceren in aangrenzende fibroblasten, waardoor wordt voorzien in de energiebehoeften van de tumorcel (Bonuccelli et al., 2010; Martinez-Outschoorn et al., 2012) . In de lever is een verschuiving in hepatocyten van ketogenese naar ketonoxidatie in hepatocellulaire carcinoom (hepatoom) cellen consistent met activering van BDH1- en SCOT-activiteiten waargenomen in twee hepatoomcellijnen (Zhang et al., 1989). Inderdaad, hepatoomcellen brengen OXCT1 en BDH1 tot expressie en oxideren ketonen, maar alleen wanneer serum uitgehongerd is (Huang et al., 2016). Als alternatief is ook ketogenese van tumorcellen voorgesteld. Dynamische verschuivingen in ketogene genexpressie worden vertoond tijdens kankertransformatie van colonepitheel, een celtype dat normaal HMGCS2 tot expressie brengt, en een recent rapport suggereerde dat HMGCS2 een prognostische marker kan zijn van een slechte prognose bij colorectale en plaveiselcelcarcinomen (Camarero et al., 2006; Chen et al., 2016). Of deze associatie ketogenese vereist of impliceert, of een maanlichtfunctie van HMGCS2, moet nog worden bepaald. Omgekeerd, schijnbare ?OHB-productie door melanoom- en glioblastoomcellen, gestimuleerd door de PPAR? agonist fenofibraat, werd geassocieerd met groeistilstand (Grabacka et al., 2016). Verdere studies zijn nodig om de rollen van HMGCS2 / SCOT-expressie, ketogenese en ketonoxidatie in kankercellen te karakteriseren.
Buiten het domein van het brandstofmetabolisme zijn ketonen recentelijk via een signaleringsmechanisme betrokken bij de biologie van kankercellen. Analyse van BRAF-V600E+ melanoom duidde op OCT1-afhankelijke inductie van HMGCL op een oncogene BRAF-afhankelijke manier (Kang et al., 2015). HMGCL-augmentatie was gecorreleerd met een hogere cellulaire AcAc-concentratie, die op zijn beurt de BRAFV600E-MEK1-interactie verbeterde, waardoor de MEK-ERK-signalering werd versterkt in een feed-forward-lus die de proliferatie en groei van tumorcellen stimuleert. Deze observaties roepen de intrigerende vraag op van prospectieve extrahepatische ketogenese die vervolgens een signaleringsmechanisme ondersteunt (zie ook ?OHB als signaleringsmediator en Controverses in extrahepatische ketogenese). Het is ook belangrijk om onafhankelijke effecten van AcAc, d-?OHB en l-?OHB op het metabolisme van kanker in overweging te nemen, en bij het overwegen van HMGCL kan leucinekatabolisme ook gestoord zijn.
De effecten van ketogene diëten (zie ook Therapeutisch gebruik van ketogeen dieet en exogene ketonlichamen) in kankerdiermodellen zijn gevarieerd (De Feyter et al., 2016; Klement et al., 2016; Meidenbauer et al., 2015; Poff et al. ., 2014; Seyfried et al., 2011; Shukla et al., 2014). Terwijl epidemiologische associaties tussen obesitas, kanker en ketogene diëten worden besproken (Liskiewicz et al., 2016; Wright en Simone, 2016), suggereerde een meta-analyse met behulp van ketogene diëten in diermodellen en in studies bij mensen een heilzaam effect op de overleving, met voordelen die prospectief zijn gekoppeld aan de omvang van ketose, het tijdstip van aanvang van het dieet en de locatie van de tumor (Klement et al., 2016; Woolf et al., 2016). Behandeling van pancreaskankercellen met ketonlichamen (d-?OHB of AcAc) remde groei, proliferatie en glycolyse, en een ketogeen dieet (81% kcal vet, 18% eiwit, 1% koolhydraat) verminderde in vivo tumorgewicht, glycemie en verhoogd spier- en lichaamsgewicht bij dieren met geïmplanteerde kanker (Shukla et al., 2014). Vergelijkbare resultaten werden waargenomen met behulp van een gemetastaseerd glioblastoomcelmodel bij muizen die ketonensuppletie in het dieet kregen (Poff et al., 2014). Omgekeerd verhoogde een ketogeen dieet (91% kcal vet, 9% eiwit) de circulerende OHB-concentratie en verminderde glycemie, maar had geen invloed op het tumorvolume of de overlevingsduur bij glioomdragende ratten (De Feyter et al., 2016). Een glucose-ketonindex is voorgesteld als een klinische indicator die de metabole behandeling van door ketogeen dieet geïnduceerde hersenkankertherapie bij mensen en muizen verbetert (Meidenbauer et al., 2015). Alles bij elkaar genomen zijn de rollen van ketonlichaammetabolisme en ketonlichamen in de kankerbiologie verleidelijk omdat ze elk handelbare therapeutische opties bieden, maar fundamentele aspecten moeten nog worden opgehelderd, met duidelijke invloeden die naar voren komen uit een matrix van variabelen, waaronder (i) verschillen tussen exogeen keton lichaam versus ketogeen dieet, (ii) kankerceltype, genomische polymorfismen, graad en stadium; en (iii) timing en duur van blootstelling aan de ketotische toestand.
Ketogenese wordt gecreëerd door ketonlichamen door de afbraak van vetzuren en ketogene aminozuren. Dit biochemische proces levert energie aan verschillende organen, met name de hersenen, onder omstandigheden van vasten als reactie op het niet beschikbaar zijn van bloedglucose. Ketonlichamen worden voornamelijk geproduceerd in de mitochondriën van levercellen. Terwijl andere cellen ketogenese kunnen uitvoeren, zijn ze daar niet zo effectief in als levercellen. Omdat ketogenese plaatsvindt in de mitochondriën, worden de processen onafhankelijk gereguleerd. Dr. Alex Jimenez DC, CCST Insight
Therapeutische toepassing van ketogeen dieet en exogene ketonlichamen
De toepassingen van ketogene diëten en ketonlichamen als therapeutische hulpmiddelen zijn ook ontstaan in niet-kankerachtige contexten, waaronder obesitas en NAFLD/NASH (Browning et al., 2011; Foster et al., 2010; Schugar en Crawford, 2012); hartfalen (Huynh, 2016; Kolwicz et al., 2016; Taegtmeyer, 2016); neurologische en neurodegeneratieve ziekte (Martin et al., 2016; McNally en Hartman, 2012; Rho, 2015; Rogawski et al., 2016; Yang en Cheng, 2010; Yao et al., 2011); aangeboren stofwisselingsziekten (Scholl-B�rgi et al, 2015); en trainingsprestaties (Cox et al., 2016). De werkzaamheid van ketogene diëten is vooral gewaardeerd bij de behandeling van epileptische aanvallen, met name bij patiënten die resistent zijn tegen geneesmiddelen. De meeste onderzoeken hebben ketogene diëten bij pediatrische patiënten geëvalueerd en laten een vermindering van de aanvalsfrequentie van ~50% zien na 3 maanden, met verbeterde effectiviteit bij bepaalde syndromen (Wu et al., 2016b). De ervaring is beperkter bij volwassen epilepsie, maar een vergelijkbare vermindering is duidelijk, met een betere respons bij symptomatische gegeneraliseerde epilepsiepatiënten (Nei et al., 2014). De onderliggende anticonvulsieve mechanismen blijven onduidelijk, hoewel gepostuleerde hypothesen een verminderd gebruik van glucose/glycolyse, geherprogrammeerd glutamaattransport, indirecte invloed op de ATP-gevoelige kaliumkanaal- of adenosine A1-receptor, wijziging van de isovormexpressie van het natriumkanaal of effecten op circulerende hormonen, waaronder leptine ( Lambrechts et al., 2016; Lin et al., 2017; Lutas en Yellen, 2013). Het blijft onduidelijk of het anti-epileptische effect voornamelijk te wijten is aan ketonlichamen, of aan de cascade metabolische gevolgen van koolhydraatarme diëten. Niettemin lijken ketonesters (zie hieronder) de aanvalsdrempel te verhogen in diermodellen van uitgelokte aanvallen (Ciarlone et al., 2016; D'Agostino et al., 2013; Viggiano et al., 2015).
Atkins-stijl en ketogene, koolhydraatarme diëten worden vaak als onaangenaam beschouwd en kunnen constipatie, hyperurikemie, hypocalciëmie, hypomagnesiëmie veroorzaken, leiden tot nefrolithiase, ketoacidose, hyperglykemie veroorzaken en de circulerende cholesterol- en vrije vetzurenconcentraties verhogen (Bisschop et al., 2001 ; Kossoff en Hartman, 2012; Kwiterovich et al., 2003; Suzuki et al., 2002). Om deze redenen vormt langdurige therapietrouw een uitdaging. Knaagdierstudies gebruiken vaak een onderscheidende verdeling van macronutriënten (94% kcal vet, 1% kcal koolhydraat, 5% kcal eiwit, Bio-Serv F3666), die een robuuste ketose veroorzaakt. Echter, het verhogen van het eiwitgehalte, zelfs tot 10% kcal, vermindert de ketose aanzienlijk, en 5% kcal eiwitbeperking geeft verwarrende metabolische en fysiologische effecten. Deze dieetformulering is ook cholinearm, een andere variabele die de gevoeligheid voor leverbeschadiging en zelfs ketogenese beïnvloedt (Garbow et al., 2011; Jornayvaz et al., 2010; Kennedy et al., 2007; Pissios et al., 2013; Schugar et al., 2013). De effecten van langdurige consumptie van ketogene diëten bij muizen blijven onvolledig gedefinieerd, maar recente studies bij muizen lieten een normale overleving en de afwezigheid van markers voor leverbeschadiging zien bij muizen op ketogene diëten gedurende hun hele leven, hoewel aminozuurmetabolisme, energieverbruik en insulinesignalering werden duidelijk opnieuw geprogrammeerd (Douris et al., 2015).
Mechanismen die ketose verhogen door mechanismen die alternatief zijn voor ketogene diëten, omvatten het gebruik van opneembare ketonlichaamprecursoren. Toediening van exogene ketonlichamen zou een unieke fysiologische toestand kunnen creëren die niet wordt aangetroffen in de normale fysiologie, omdat de circulerende glucose- en insulineconcentraties relatief normaal zijn, terwijl cellen de opname en het gebruik van glucose kunnen sparen. Ketonlichamen zelf hebben korte halfwaardetijden en inname of infusie van natrium-OHB-zout om therapeutische ketose te bereiken veroorzaakt een ongewenste natriumbelasting. R/S-1,3-butaandiol is een niet-toxische dialcohol die gemakkelijk in de lever wordt geoxideerd om d/l-?OHB op te leveren (Desrochers et al., 1992). In verschillende experimentele contexten is deze dosis gedurende zeven weken dagelijks aan muizen of ratten toegediend, wat circulerende ?OHB-concentraties opleverde van maximaal 5 mM binnen 2 uur na toediening, die stabiel is gedurende ten minste nog eens 3 uur (D' Agostino et al., 2013). Gedeeltelijke onderdrukking van de voedselinname is waargenomen bij knaagdieren die R/S-1,3-butaandiol kregen (Carpenter en Grossman, 1983). Daarnaast drie chemisch verschillende keton esters (KE's), (i) monoester van R-1,3-butaandiol en d-?OHB (R-3-hydroxybutyl R-?OHB); (ii) glyceryl-tris-?OHB; en (iii) R,S-1,3-butaandiol-acetoacetaat-diester zijn ook uitgebreid bestudeerd (Brunengraber, 1997; Clarke et al., 2012a; Clarke et al., 2012b; Desrochers et al., 1995a; Desrochers et al. ., 1995b; Kashiwaya et al., 2010). Een inherent voordeel van de eerste is dat er 2 mol fysiologisch d-?OHB wordt geproduceerd per mol KE, na hydrolyse van esterase in de darm of lever. Veiligheid, farmacokinetiek en tolerantie zijn het meest uitgebreid bestudeerd bij mensen die R-3-hydroxybutyl R-?OHB innamen, in doses tot 714 mg/kg, wat circulerende d-?OHB-concentraties tot 6 mM opleverde (Clarke et al., 2012a; Cox et al., 2016; Kemper et al., 2015; Shivva et al., 2016). Bij knaagdieren verlaagt deze KE de calorie-inname en het totale cholesterolgehalte in het plasma, stimuleert het bruin vetweefsel en verbetert het de insulineresistentie (Kashiwaya et al., 2010; Kemper et al., 2015; Veech, 2013). Recente bevindingen geven aan dat tijdens inspanning bij getrainde atleten de inname van R-3-hydroxybutyl R-?OHB de glycolyse van de skeletspieren en de plasmalactaatconcentraties verminderde, de intramusculaire triacylglyceroloxidatie verhoogde en het spierglycogeengehalte in stand hield, zelfs wanneer gelijktijdige inname van koolhydraten de insulinesecretie stimuleerde ( Cox et al., 2016). Verdere ontwikkeling van deze intrigerende resultaten is vereist, omdat de verbetering van de prestaties van de duurtraining voornamelijk werd veroorzaakt door een robuuste respons op de KE bij 2/8 proefpersonen. Desalniettemin ondersteunen deze resultaten klassieke onderzoeken die wijzen op een voorkeur voor ketonoxidatie boven andere substraten (GARLAND et al., 1962; Hasselbaink et al., 2003; Stanley et al., 2003; Valente-Silva et al., 2015), ook tijdens inspanning, en dat getrainde atleten meer geneigd zijn om ketonen te gebruiken (Johnson et al., 1969a; Johnson en Walton, 1972; Winder et al., 1974; Winder et al., 1975). Ten slotte moeten de mechanismen die verbeterde trainingsprestaties kunnen ondersteunen na gelijke calorie-inname (differentieel verdeeld over macronutriënten) en gelijke zuurstofconsumptie nog worden bepaald.
Toekomstperspectief
Eens grotendeels gestigmatiseerd als een overlooproute die in staat is om giftige emissies van vetverbranding te accumuleren in staten met beperkte koolhydraten (het 'ketotoxische' paradigma), ondersteunen recente waarnemingen het idee dat het ketonlichaammetabolisme een heilzame rol speelt, zelfs in met koolhydraten beladen toestanden. � hypothese. Hoewel de gemakkelijke nutritionele en farmacologische benaderingen om ketonmetabolisme te manipuleren het een aantrekkelijk therapeutisch doelwit maken, blijven agressief geponeerde maar voorzichtige experimenten in zowel de basis- als translationele onderzoekslaboratoria. Er zijn onvervulde behoeften ontstaan op het gebied van het definiëren van de rol van het gebruik van ketonmetabolisme bij hartfalen, obesitas, NAFLD/NASH, diabetes type 2 en kanker. De reikwijdte en impact van 'niet-canonieke' signaleringsrollen van ketonlichamen, inclusief regulatie van PTM's die waarschijnlijk heen en weer terugkoppelen naar metabole en signaalroutes, vereisen een diepere verkenning. Ten slotte zou extrahepatische ketogenese intrigerende paracriene en autocriene signaleringsmechanismen en mogelijkheden kunnen openen om het co-metabolisme in het zenuwstelsel en tumoren te beïnvloeden om therapeutische doeleinden te bereiken.
Concluderend, ketonlichamen worden door de lever aangemaakt om als energiebron te worden gebruikt wanneer er niet genoeg glucose direct beschikbaar is in het menselijk lichaam. Ketogenese treedt op wanneer er lage glucosespiegels in het bloed zijn, vooral nadat andere cellulaire koolhydraatvoorraden zijn uitgeput. Het doel van het bovenstaande artikel was om de multidimensionale rol van ketonlichamen in brandstofmetabolisme, signalering en therapieën te bespreken. De reikwijdte van onze informatie is beperkt tot chiropractische en spinale gezondheidsproblemen. Als u het onderwerp wilt bespreken, kunt u het aan Dr. Jimenez vragen of contact met ons opnemen via�915-850-0900 .
Rugpijn is een van de meest voorkomende oorzaken van arbeidsongeschiktheid en gemiste werkdagen wereldwijd. Rugpijn is de tweede meest voorkomende reden voor doktersbezoeken, alleen overtroffen door bovenste luchtweginfecties. Ongeveer 80 procent van de bevolking zal gedurende zijn leven minstens één keer rugpijn ervaren. De wervelkolom is een complexe structuur die bestaat uit botten, gewrichten, ligamenten en spieren, naast andere zachte weefsels. Letsel en / of verergerde omstandigheden, zoals herniated discs, kan uiteindelijk leiden tot symptomen van rugpijn. Sportblessures of auto-ongelukken zijn vaak de meest voorkomende oorzaak van rugpijn, maar soms kunnen de eenvoudigste bewegingen pijnlijke gevolgen hebben. Gelukkig kunnen alternatieve behandelingsopties, zoals chiropractische zorg, rugpijn helpen verlichten door het gebruik van spinale aanpassingen en handmatige manipulaties, waardoor uiteindelijk de pijnverlichting wordt verbeterd. �
De nucleaire erytroïde 2-gerelateerde factor 2-signaleringsroute, beter bekend als NrF2, is een beschermend mechanisme dat fungeert als een 'hoofdregelaar' van de antioxidantrespons van het menselijk lichaam. Nrf2 detecteert de niveaus van oxidatieve stress in de cellen en veroorzaakt beschermende antioxidante mechanismen. Hoewel Nrf2-activering vele voordelen kan hebben, kan Nrf2 "overexpressie" verschillende risico's met zich meebrengen.
Het lijkt erop dat een evenwichtige mate van NRF2 essentieel is om de algehele ontwikkeling van een verscheidenheid aan ziekten te voorkomen, naast de algemene verbetering van deze gezondheidskwesties. NRF2 kan echter ook complicaties veroorzaken. De belangrijkste oorzaak achter NRF2 "overexpressie" is te wijten aan een genetische mutatie of een aanhoudende chronische blootstelling aan onder andere een chemische of oxidatieve stress. Hieronder bespreken we de nadelen van overfxpressie van Nrf2 en demonstreren we de werkingsmechanismen ervan in het menselijk lichaam.
Kanker
Onderzoeksstudies hebben aangetoond dat muizen die NRF2 niet tot expressie brengen, meer geneigd zijn om kanker te ontwikkelen als reactie op fysieke en chemische stimulatie. Vergelijkbare onderzoeken hebben echter aangetoond dat overactivering van NRF2, of zelfs inactivering van KEAP1, kan leiden tot verergering van bepaalde kankers, vooral als die paden zijn onderbroken. Overactief NRF2 kan optreden door roken, waarbij wordt aangenomen dat continue NRF2-activering de oorzaak is van longkanker bij rokers. Overexpressie van Nrf2 kan ervoor zorgen dat kankercellen zichzelf niet vernietigen, terwijl intermitterende NRF2-activering kan voorkomen dat kankercellen toxine-inductie veroorzaken.
Omdat overexpressie van NRF2 het antioxiderende vermogen van het menselijk lichaam om verder te gaan dan redoxhomeostase verhoogt, stimuleert dit bovendien de celdeling en genereert het een onnatuurlijk patroon van DNA- en histonmethylering. Dit kan uiteindelijk chemotherapie en radiotherapie minder effectief maken tegen kanker. Daarom zou het beperken van NRF2-activering met stoffen zoals DIM, Luteoline, Zi Cao of salinomycine ideaal kunnen zijn voor patiënten met kanker, hoewel overactivering van Nrf2 niet als de enige oorzaak van kanker moet worden beschouwd. Tekorten aan voedingsstoffen kunnen genen aantasten, waaronder NRF2. Dit kan een manier zijn om te zien hoe tekortkomingen bijdragen aan tumoren.
Lever
De overactivering van Nrf2 kan ook de functie van specifieke organen in het menselijk lichaam beïnvloeden. NRF2-overexpressie kan uiteindelijk de productie van de insuline-achtige groeifactor 1 of IGF-1 uit de lever blokkeren, wat essentieel is voor de regeneratie van de lever.
Hart
Hoewel de acute overexpressie van Nrf2 de voordelen kan hebben, kan voortdurende overexpressie van NRF2 op lange termijn schadelijke effecten op het hart veroorzaken, zoals cardiomyopathie. NRF2-expressie kan worden verhoogd door hoge niveaus van cholesterol of de activering van HO-1. Dit wordt verondersteld de reden te zijn waarom chronische verhoogde niveaus van cholesterol cardiovasculaire gezondheidsproblemen kunnen veroorzaken.
Vitiligo
NRF2-overexpressie heeft ook aangetoond dat het de mogelijkheid van repigment in vitiligo remt, omdat het Tyrosinase of TYR, een actie die essentieel is voor repigmentatie door melaninogenese, kan belemmeren. Onderzoeksstudies hebben aangetoond dat dit proces een van de belangrijkste redenen kan zijn waarom mensen met vitiligo NRF2 niet zo efficiënt lijken te activeren als mensen zonder vitiligo.
Waarom NRF2 misschien niet goed functioneert
hormese
NRF2 moet hormetisch worden geactiveerd om te kunnen profiteren van de voordelen ervan. Met andere woorden, Nrf2 mag niet elke minuut of elke dag worden geactiveerd, daarom is het een goed idee om er een pauze van te nemen, bijvoorbeeld 5 dagen op 5 vrije dagen of om de dag. NRF2 moet ook een specifieke drempel bereiken om zijn hormetische respons te activeren, waar een kleine stressfactor mogelijk niet voldoende is om het te activeren.
DJ-1 Oxidatie
Protein deglycase DJ-1, of gewoon DJ-1, ook wel het Parkinson-eiwit genoemd, of PARK7, is een hoofdregelaar en detector van de redox-status in het menselijk lichaam. DJ-1 is essentieel om te regelen hoelang NRF2 zijn functie kan vervullen en een antioxiderend effect kan produceren. In het geval dat DJ-1 overgeoxideerd raakt, maken de cellen het DJ-1-eiwit minder toegankelijk.
Dit proces zorgt ervoor dat de NRF2-activering te snel verloopt, omdat DJ-1 van het grootste belang is om de gebalanceerde niveaus van NRF2 te behouden en te voorkomen dat ze worden afgebroken in de cel. Als het DJ-1-eiwit niet bestaat of overgeoxideerd is, zal NRF2-expressie waarschijnlijk minimaal zijn, zelfs met DIM of alternatieve NRF2-activators. DJ-1-expressie is noodzakelijk om een gestoorde NRF2-actie te herstellen.
Chronische Ziekte
Als u een chronische ziekte heeft, waaronder CIRS, chronische infecties/dysbiose/SIBO, of ophoping van zware metalen, zoals kwik en/of die van wortelkanalen, kunnen deze de systemen van NRF2 en fase twee ontgifting belemmeren. In plaats van oxidatieve stress die NRF2 in een antioxidant verandert, zal NRF2 niet triggeren en oxidatieve stress kan in de cel blijven en schade veroorzaken, wat betekent dat er geen antioxidantreactie is. Dit is een belangrijke reden waarom veel mensen met CIRS verschillende gevoeligheden hebben en veel factoren bereiken. Sommige mensen denken dat ze een herxreactie hebben, maar deze reactie kan de cellen alleen maar verder beschadigen.
Door chronische ziekten te behandelen, kan de lever echter gifstoffen in de gal afvoeren, waardoor de hormetische respons van NRF2-activering geleidelijk wordt ontwikkeld. Als de gal giftig blijft en niet uit het menselijk lichaam wordt uitgescheiden, zal het de oxidatieve stress van NRF2 reactiveren en ervoor zorgen dat u zich slechter gaat voelen zodra het opnieuw is opgenomen uit het maagdarmkanaal of het maagdarmkanaal. Ochratoxine A kan bijvoorbeeld NRF2 blokkeren. Afgezien van het behandelen van het probleem, kunnen histondeacetylaseremmers de oxidatieve reactie van een aantal van de factoren die NRF2-activering veroorzaken, blokkeren, maar het kan ook voorkomen dat NRF2 normaal triggert, wat uiteindelijk zijn doel niet zou kunnen dienen.
Visolie ontregeling
Cholinergica zijn stoffen die acetylcholine, oftewel ACh, en choline in de hersenen stimuleren door de toename van ACh, met name bij het remmen van de afbraak van ACh. Patiënten met CIRS hebben vaak problemen met de ontregeling van acetylcholinegehaltes in het menselijk lichaam, vooral in de hersenen. Visolie triggert NRF2 en activeert het beschermende antioxidantmechanisme in de cellen.
Mensen met chronische ziekten kunnen problemen hebben met cognitieve stress en acetylcholine-excitotoxiciteit, door ophoping van organofosfaat, waardoor visolie een ontsteking in het menselijk lichaam kan veroorzaken. Choline-deficiëntie induceert bovendien NRF2-activering. Het opnemen van choline in uw dieet (polyfenolen, eieren, enz.) kan de effecten van cholinerge ontregeling helpen versterken.
Wat vermindert NRF2?
Een afname van NRF2-overexpressie is het beste voor mensen die kanker hebben, hoewel dit voor veel andere gezondheidsproblemen van pas kan komen.
Dieet, supplementen en gemeenschappelijke medicijnen:
Apigenin (hogere doses)
Brucea javanica
kastanjes
EGCG (hoge doses verhogen NRF2)
Fenegriek (Trigonelline)
Hiba (Hinokitiol /? -Thujaplicine)
Hoog zoutdieet
Luteolin (selderij, groene paprika, peterselie, perilla-blad en kamille-thee - hogere doses kunnen NRF2 verhogen - 40 mg / kg luteoline driemaal per week)
Metformine (chronische inname)
N-Acetyl-L-Cysteïne (NAC, door de oxidatieve reactie te blokkeren, vooral in hoge doses)
Quercetine (hogere doses kunnen NRF2 verhogen - 50 mg / kg / d quercetine)
Salinomycin (drug)
Retinol (volledig trans-retinezuur)
Vitamine C in combinatie met Quercetine
Zi Cao (Purple Gromwel heeft Shikonin / Alkannin)
Pathways and Other:
Bach1
BET
biofilms
Brusatol
camptothecine
DNMT
DPP-23
EZH2
Glucocorticoïd Receptor-signalering (ook Dexamethason en Betamethason)
GSK-3? (regelgevende feedback)
HDAC-activering?
Halofuginon
Homocysteïne (ALCAR kan deze homocysteïne omkeren, induceert lage niveaus van NRF2)
IL-24
Keap1
MDA-7
NF? B
Ochratoxine A (aspergillus en pencicllium soorten)
Promyelocytisch leukemie-eiwit
p38
p53
p97
Retinoïnezuurreceptor alfa
Seleniet
SYVN1 (Hrd1)
STAT3-remming (zoals Cryptotanshinone)
Testosteron (en testosteronpropionaat, hoewel TP intranasaal NRF2 kan verhogen)
Trecator (Ethionamide)
Trx1 (via reductie van Cys151 in Keap1 of van Cys506 in de NLS-regio van Nrf2)
Trolox
Vorinostat
Zinc Deficiency (maakt het erger in de hersenen)
Nrf2 Werkingsmechanisme
Oxidatieve stress wordt geactiveerd via CUL3, waarbij NRF2 van KEAP1, een negatieve remmer, vervolgens de kern van deze cellen binnengaat, de transcriptie van de ARE's stimuleert, sulfiden omzet in disulfiden en ze verandert in meer antioxidante genen, wat leidt tot de opregulering van antioxidanten, zoals als GSH, GPX, GST, SOD, enz. De rest van deze zijn te zien in de onderstaande lijst:
Verhoogt AKR
Verhoogt ZE
Verhoogt ATF4
Verhoogt Bcl-xL
Verhoogt Bcl-2
Verhoogt BDNF
Verhoogt BRCA1
Verhoogt c-Jun
Vergroot CAT
Verhoogt cGMP
Verhoogt CKIP-1
Verhoogt CYP450
Verhoogt Cul3
Verhoogt GCL
Verhoogt GCLC
Verhoogt GCLM
Verhoogt GCS
Verhoogt GPx
Verhoogt GR
Verhoogt GSH
Verhoogt GST
Verhoogt HIF1
Verhoogt HO-1
Verhoogt HQO1
Verhoogt HSP70
Verhoogt IL-4
Verhoogt IL-5
Verhoogt IL-10
Verhoogt IL-13
Verhoogt K6
Verhoogt K16
Verhoogt K17
Verhoogt mEH
Verhoogt Mrp2-5
Verhoogt NADPH
Vergroot Notch 1
Verhoogt NQO1
Verhoogt PPAR-alpha
Verhoogt Prx
Verhoogt p62
Verhoogt Sesn2
Verhoogt Slco1b2
Verhoogt sMafs
Verhoogt SOD
Verhoogt Trx
Verhoogt Txn (d)
Verhoogt UGT1 (A1 / 6)
Verhoogt VEGF
Vermindert ADAMTS (4 / 5)
Vermindert alpha-SMA
Vermindert ALT
Vermindert AP1
Vermindert AST
Vermindert Bach1
Vermindert COX-2
Vermindert DNMT
Vermindert FASN
Reduceert FGF
Reduceert HDAC
Vermindert IFN-?
Vermindert IgE
Vermindert IGF-1
Vermindert IL-1b
Vermindert IL-2
Vermindert IL-6
Vermindert IL-8
Vermindert IL-25
Vermindert IL-33
Vermindert iNOS
Reduceert LT
Reduceert Keap1
Reduceert MCP-1
Reduceert MIP-2
Reduceert MMP-1
Reduceert MMP-2
Reduceert MMP-3
Reduceert MMP-9
Reduceert MMP-13
Reduceert NfkB
Reduceert NO
Reduceert SIRT1
Vermindert TGF-b1
Vermindert TNF-alfa
Reduceert Tyr
Reduceert VCAM-1
Gecodeerd van het NFE2L2-gen is NRF2 of kern-erytroïde 2-gerelateerde factor 2 een transcriptiefactor in de basische leucineritssluiting of bZIP, superfamilie die een Cap'n'Collar- of CNC-structuur gebruikt.
Het bevordert salpeterzymen, biotransformatie-enzymen en xenobiotische effluxtransporters.
Het is een essentiële regulator bij de inductie van de fase II-genen voor antioxidanten en ontgiftingsenzymen, die cellen beschermen tegen schade veroorzaakt door oxidatieve stress en elektrofiele aanvallen.
Tijdens homeostatische aandoeningen wordt Nrf2 gesekwestreerd in het cytosol door middel van lichamelijke hechting van het N-terminale domein van Nrf2, of het Kelch-achtige ECH-geassocieerde eiwit of Keap1, ook wel INrf2 of Inhibitor van Nrf2 genoemd, remt de Nrf2-activering.
Het kan ook worden gereguleerd door zoogdier-selenoproteïne thioredoxine-reductase 1 of TrxR1, dat als een negatieve regulator fungeert.
Bij kwetsbaarheid voor elektrofiele stressfactoren dissocieert Nrf2 van Keap1, translocatie in de kern, waar het vervolgens heterodimeriseert met een reeks transcriptionele regulatoire eiwitten.
Frequente interacties omvatten met die van transcriptie-autoriteiten Jun en Fos, die lid kunnen zijn van de activatorproteïnefamilie van transcriptiefactoren.
Na dimerisatie binden deze complexen zich vervolgens aan antioxidant / elektrofiel reagerende componenten ARE / EpRE en activeren ze transcriptie, zoals waar is met het Jun-Nrf2-complex, of onderdrukken ze transcriptie, vergelijkbaar met het Fos-Nrf2-complex.
De positionering van de ARE, die wordt geactiveerd of geremd, zal bepalen welke genen transcriptioneel worden gecontroleerd door deze variabelen.
Wanneer ARE wordt geactiveerd:
Activering van de synthese van antioxidanten is in staat om ROS te ontgiften zoals catalase, superoxide-dismutase of SOD, GSH-peroxidasen, GSH-reductase, GSH-transferase, NADPH-chinonoxidoreductase of NQO1, Cytochrome P450 mono-oxygenasesysteem, thioredoxine, thioredoxine reductase en HSP70.
Activering van dit GSH-synthase maakt een merkbare groei van de intracellulaire GSH-graad mogelijk, wat behoorlijk beschermend is.
De toename van deze synthese en gradaties van fase II-enzymen zoals UDP-glucuronosyltransferase, N-acetyltransferasen en sulfotransferasen.
De opregulatie van HO-1, wat een echt beschermende receptor is met een potentiële CO-groei die in combinatie met NO vasodilatatie van ischemische cellen mogelijk maakt.
Vermindering van ijzerstapeling door verhoogd ferritine en bilirubine als lipofiel antioxidant. Zowel de fase II-eiwitten als de antioxidanten zijn in staat om de chronische oxidatieve stress te herstellen en ook om een normaal redoxsysteem te doen herleven.
GSK3? onder het beheer van AKT en PI3K, fosforyleert Fyn resulterend in Fyn nucleaire lokalisatie, die Fyn fosforyleert Nrf2Y568 leidend tot nucleaire export en afbraak van Nrf2.
NRF2 dempt ook de TH1 / TH17-respons en verrijkt de TH2-respons.
HDAC-remmers activeerden de Nrf2-signaleringsroute en omhoog gereguleerd dat de Nrf2 stroomafwaarts HO-1, NQO1 en glutamaat-cysteïne ligase katalytische subeenheid, of GCLC, target door Keap1 te beperken en de dissociatie van Keap1 van Nrf2, Nrf2 nucleaire translocatie en Nrf2 aan te moedigen -ARE bindend.
Nrf2 bevat een halfwaardetijd van ongeveer 20 minuten onder basale omstandigheden.
De IKK verminderen? pool via Keap1-binding vermindert I? B? afbraak en zou het ongrijpbare mechanisme kunnen zijn waarmee is bewezen dat Nrf2-activering NF? B-activering remt.
Keap1 hoeft niet altijd te worden gereguleerd om NRF2 te laten werken, zoals chlorophyllin, blueberry, ellagic acid, astaxanthin en tea polyphenols kunnen NRF2 en KEAP1 met 400 procent stimuleren.
Nrf2 reguleert negatief door de duur van stearoyl CoA-desaturase, of SCD, en citraatlyase, of CL.
Genetica
KEAP1
rs1048290
C-allel - vertoonde een aanzienlijk risico voor en een beschermend effect tegen geneesmiddelresistente epilepsie (DRE)
rs11085735 (ik ben AC)
geassocieerd met snelheid van verval van longfunctie in de LHS
MAPT
rs242561
T-allel - beschermend allel voor Parkinsonstoornissen - had sterkere NRF2 / sMAF-binding en was geassocieerd met de hogere MAPT-mRNA-niveaus in 3-verschillende regio's in de hersenen, inclusief cerebellaire cortex (CRBL), temporale cortex (TCTX), intralobulaire witte stof (WHMT)
NFE2L2 (NRF2)
rs10183914 (ik ben CT)
T-allel - verhoogde niveaus van Nrf2-eiwit en vertraagde leeftijd bij het optreden van Parkinson met vier jaar
rs16865105 (ik ben AC)
C-allel - had een hoger risico op de ziekte van Parkinson
rs1806649 (ik ben CT)
C-allel - is geïdentificeerd en kan relevant zijn voor de etiologie van borstkanker.
geassocieerd met een verhoogd risico op ziekenhuisopnamen tijdens periodes met hoge PM10-niveaus
rs1962142 (ik ben GG)
T-allel - was geassocieerd met een laag niveau van cytoplasmatische NRF2-expressie (P = 0.036) en negatieve sulfiredoxine-expressie (P = 0.042)
Een allel - beschermd tegen onderarmbloedstroom (FEV) -afname (geforceerd expiratoir volume in één seconde) in relatie tot de status van het roken van sigaretten (p = 0.004)
rs2001350 (ik ben TT)
T-allel - beschermd tegen FEV-afname (geforceerd expiratoir volume in één seconde) met betrekking tot de status van het roken van sigaretten (p = 0.004)
rs2364722 (ik ben AA)
Een allel - beschermd tegen FEV-afname (geforceerd expiratoir volume in één seconde) met betrekking tot de status van het roken van sigaretten (p = 0.004)
rs2364723
C-allel - geassocieerd met aanzienlijk verminderde FEV bij Japanse rokers met longkanker
rs2706110
G-allel - toonde een significant risico voor en een beschermend effect tegen geneesmiddelresistente epilepsie (DRE)
AA-allelen - was geassocieerd met een verhoogd risico op borstkanker (P = 0.011)
rs2886161 (ik ben TT)
T-allel - geassocieerd met de ziekte van Parkinson
rs2886162
Een allel - was geassocieerd met lage NRF2-expressie (P = 0.011; OR, 1.988; CI, 1.162) en het AA-genotype was geassocieerd met een slechtere overleving (P = 3.400; HR, 0.032; CI, 1.687-1.047)
rs35652124 (ik ben TT)
Een allel - geassocieerd met hoger geassocieerd met leeftijd bij aanvang van het Parkinson-ziekte versus G-allel
C-allel - verhoogde NRF2-eiwit
T-allel - had minder NRF2-eiwit en een groter risico op hartaandoeningen en bloeddruk
rs6706649 (ik ben CC)
C-allel - had lager NRF2-eiwit en verhoogt het risico op de ziekte van Parkinson
rs6721961 (ik ben GG)
T-allel - had lager NRF2-eiwit
TT-allelen - verband tussen het roken van sigaretten bij zware rokers en een afname van de spermakwaliteit
TT-allel – ging gepaard met een verhoogd risico op borstkanker [P = 0.008; OF, 4.656; betrouwbaarheidsinterval (BI), 1.350 - 16.063] en het T-allel was geassocieerd met een lage mate van NRF2-eiwitexpressie (P = 0.0003; OR, 2.420; CI, 1.491 - 3.926) en negatieve SRXN1-expressie (P = 0.047; OR, 1.867; BI = 1.002-3.478)
T-allel - allel was ook nominaal geassocieerd met ALI-gerelateerde 28-dag mortaliteit na systemisch inflammatoir respons syndroom
T-allel - beschermd tegen FEV-afname (geforceerd expiratoir volume in één seconde) met betrekking tot de status van het roken van sigaretten (p = 0.004)
G-allel - geassocieerd met een verhoogd risico op ALI na een groot trauma bij Europese en Afro-Amerikanen (odds ratio, OR 6.44; 95% betrouwbaarheidsinterval
AA-allelen - geassocieerd met door infectie geïnduceerde astma
AA-allelen - vertoonde significant verminderde NRF2-genexpressie en bijgevolg een verhoogd risico op longkanker, vooral degenen die ooit hadden gerookt
AA-allelen - hadden een significant hoger risico voor de ontwikkeling van T2DM (OF 1.77; 95% CI 1.26, 2.49; p = 0.011) in verhouding tot die met het CC-genotype
AA-allelen - sterke associatie tussen wondherstel en late toxiciteiten van straling (geassocieerd met een significant hoger risico op het ontwikkelen van late effecten bij Afro-Amerikanen met een trend in blanken)
geassocieerd met orale oestrogeentherapie en risico op veneuze trombo-embolie bij postmenopauzale vrouwen
rs6726395 (ik ben AG)
Een allel - beschermd tegen FEV1-afname (geforceerd expiratoir volume in één seconde) met betrekking tot de status van het roken van sigaretten (p = 0.004)
Een allel - geassocieerd met aanzienlijk verminderde FEV1 bij Japanse rokers met longkanker
GG-allelen - hadden hogere NRF2-niveaus en verminderden het risico op maculaire degeneratie
GG-allelen - hadden een hogere overleving met Cholangiocarcinoom
rs7557529 (ik ben CT)
C-allel - geassocieerd met de ziekte van Parkinson
Oxidatieve stress en andere stressfactoren kunnen celbeschadiging veroorzaken die uiteindelijk tot een verscheidenheid aan gezondheidsproblemen kan leiden. Onderzoeksstudies hebben aangetoond dat Nrf2-activering het beschermende antioxidantmechanisme van het menselijk lichaam kan bevorderen, maar onderzoekers hebben besproken dat overexpressie van Nrf2 enorme risico's met zich mee kan brengen voor de algehele gezondheid en het welzijn. Verschillende soorten kanker kunnen ook optreden met overfactivering van Nrf2.
Dr. Alex Jimenez DC, CCST Insight
Sulforafaan en de effecten ervan op kanker, mortaliteit, veroudering, hersenen en gedrag, hartaandoeningen en meer
Isothiocyanaten zijn enkele van de belangrijkste plantaardige stoffen die u in uw dieet kunt krijgen. In deze video maak ik de meest uitgebreide case voor hen die ooit is gemaakt. Korte aandachtsspanne? Ga naar uw favoriete onderwerp door op een van de onderstaande tijdspunten te klikken. Volledige tijdlijn hieronder.
Belangrijkste secties:
00: 01: 14 - Kanker en sterfte
00: 19: 04 - Veroudering
00: 26: 30 - Hersenen en gedrag
00: 38: 06 - Laatste overzicht
00: 40: 27 - Dosis
Volledige tijdlijn:
00: 00: 34 - Introductie van sulforafaan, een belangrijk aandachtspunt van de video.
00: 01: 14 - Kruisbloemige groenteconsumptie en verlagingen van mortaliteit door alle oorzaken.
00: 02: 12 - Risico op prostaatkanker.
00: 02: 23 - Blaaskankerrisico.
00: 02: 34 - Longkanker bij rokers risico.
00: 02: 48 - Risico op borstkanker.
00: 03: 13 - Hypothetisch: wat als je al kanker hebt? (Interventie)
00: 03: 35 - Aannemelijk mechanisme dat de associatieve gegevens over kanker en mortaliteit stuurt.
00: 04: 38 - Sulforafaan en kanker.
00: 05: 32 - Dierlijk bewijs dat een sterk effect van extract van broccolispruit toont op de ontwikkeling van blaastumoren bij ratten.
00: 06: 06 - Effect van directe suppletie van sulforafaan bij prostaatkankerpatiënten.
00: 07: 09 - Bioaccumulatie van isothiocyanaatmetabolieten in daadwerkelijk borstweefsel.
00: 08: 32 - Remming van stamcellen van borstkanker.
00: 08: 53 - Geschiedenisles: brassica's werden vastgesteld met gezondheidseigenschappen zelfs in het oude Rome.
00: 09: 16 - het vermogen van Sulforaphane om de kankerverwekkende uitscheiding te verbeteren (benzeen, acroleïne).
00: 09: 51 - NRF2 als een genetische switch via antioxidantresponselementen.
00: 10: 10 - Hoe NRF2-activering de carcinogene excretie via glutathion-S-conjugaten verbetert.
00: 10: 34 - Spruitjes verhogen glutathion-S-transferase en verminderen DNA-schade.
00: 11: 20 - Broccolispruitendrank verhoogt de benzeenuitscheiding met 61%.
00: 13: 31 - Broccoli-spruithomogenaat verhoogt antioxiderende enzymen in de bovenste luchtwegen.
00: 15: 45 - Kruisbloemige groenteconsumptie en sterfte aan hart- en vaatziekten.
00: 16: 55 - Broccolispruitpoeder verbetert de bloedlipiden en het algemene risico op hartziekten bij type 2 diabetici.
00: 19: 04 - Begin van het verouderingsgedeelte.
00: 19: 21 - Met Sulforaphane verrijkte voeding verlengt de levensduur van kevers van 15 tot 30% (onder bepaalde omstandigheden).
00: 20: 34 - Belang van lage ontsteking voor een lang leven.
00: 22: 05 - Kruisbloemige groenten en broccoli-kiempoeder lijken een breed scala aan ontstekingsmarkers bij mensen te verminderen.
00: 24: 14 - Muisstudies suggereren dat sulforaphane de adaptieve immuunfunctie op oudere leeftijd zou kunnen verbeteren.
00: 25: 18 - Sulforafaan verbeterde de haargroei in een muismodel van kalend. Afbeelding bij 00: 26: 10.
00: 26: 30 - Begin van de sectie hersenen en gedrag.
00: 27: 18 - Effect van extract van broccolispruiten op autisme.
00: 27: 48 - Effect van glucoraphanine op schizofrenie.
00: 28: 17 - Begin van depressie discussie (plausibel mechanisme en studies).
00: 31: 21 - Muisstudie met behulp van 10 verschillende modellen van stress-geïnduceerde depressie laten sulforafaan zien, even effectief als fluoxetine (prozac).
00: 32: 00 - Onderzoek toont aan dat directe inname van glucoraphanine bij muizen even effectief is bij het voorkomen van depressies als gevolg van een sociaal nederlaagstressmodel.
00: 33: 01 - Begin van de sectie neurodegeneratie.
00: 33: 30 - Sulforafane en de ziekte van Alzheimer.
00: 33: 44 - Sulforafane en de ziekte van Parkinson.
00: 33: 51 - De ziekte van Sulforaphane en Hungtington.
00: 34: 43 - Begin van een traumatisch gedeelte over hersenletsel.
00: 35: 01 - Sulforafaan wordt geïnjecteerd onmiddellijk nadat TBI het geheugen verbetert (muisstudie).
00: 35: 55 - Sulforafaan en neuronale plasticiteit.
00: 36: 32 - Sulforafaan verbetert het leren in het model van type II diabetes bij muizen.
00: 37: 19 - Sulforafaan en duchenne spierdystrofie.
00: 37: 44 - Myostatinremming in spieratlanticellen (in vitro).
00: 38: 06 - Late video recapitulatie: sterfte en kanker, DNA-schade, oxidatieve stress en ontsteking, uitscheiding van benzeen, cardiovasculaire aandoeningen, type II diabetes, effecten op de hersenen (depressie, autisme, schizofrenie, neurodegeneratie), NRF2-route.
00: 40: 27 - Gedachten over het uitzoeken van een dosis broccoli of sulforafaan.
00: 41: 01 - Anekdotes over ontspruiten thuis.
00: 43: 14 - Op kooktemperaturen en sulforafaanactiviteit.
00: 43: 45 - Darmbacterieomzetting van sulforafaan uit glucoraphanine.
00: 44: 24 - Supplementen werken beter in combinatie met actieve myrosinase van groenten.
00: 44: 56 - Koken technieken en kruisbloemige groenten.
00: 46: 06 - Isothiocyanaten als goitrogens.
Volgens onderzoeksstudies is Nrf2 een fundamentele transcriptiefactor die de beschermende antioxidantmechanismen van de cellen activeert om het menselijk lichaam te ontgiften. De overexpressie van Nrf2 kan echter gezondheidsproblemen veroorzaken. De reikwijdte van onze informatie is beperkt tot chiropractische en spinale gezondheidsproblemen. Als u het onderwerp wilt bespreken, kunt u het aan Dr. Jimenez vragen of contact met ons opnemen via�915-850-0900 .
Curator van Dr. Alex Jimenez
Bijkomend onderwerpbespreking: Acute rugpijn
Rugpijn is een van de meest voorkomende oorzaken van arbeidsongeschiktheid en gemiste werkdagen wereldwijd. Rugpijn is de tweede meest voorkomende reden voor doktersbezoeken, alleen overtroffen door bovenste luchtweginfecties. Ongeveer 80 procent van de bevolking zal gedurende zijn leven minstens één keer rugpijn ervaren. De wervelkolom is een complexe structuur die bestaat uit botten, gewrichten, ligamenten en spieren, naast andere zachte weefsels. Letsel en / of verergerde omstandigheden, zoals herniated discs, kan uiteindelijk leiden tot symptomen van rugpijn. Sportblessures of ongevallen met auto-ongelukken zijn vaak de meest voorkomende oorzaak van rugpijn, maar soms kunnen de eenvoudigste bewegingen pijnlijke resultaten hebben. Gelukkig kunnen alternatieve behandelingsopties, zoals chiropractische zorg, de rugpijn verlichten door het gebruik van spinale aanpassingen en handmatige manipulaties, waardoor uiteindelijk de pijnverlichting wordt verbeterd.
Veel van de huidige onderzoeken naar kanker hebben gezondheidswerkers in staat gesteld te begrijpen hoe het lichaam detoxeert. Door upregulated genen in tumorcellen te analyseren, ontdekten onderzoekers de nucleaire erytroïde 2-gerelateerde factor 2-signaleringsroute, het best bekend als Nrf2. NRF2 is een belangrijke transcriptiefactor die het menselijk lichaam activeert beschermende antioxidante mechanismen om de oxidatie van zowel externe als interne factoren te reguleren om verhoogde niveaus van oxidatieve stress te voorkomen.
Principes van Nrf2
NRF2 is essentieel voor het handhaven van de algehele gezondheid en welzijn, omdat het het primaire doel dient om te reguleren hoe we omgaan met alles waaraan we dagelijks worden blootgesteld en niet ziek worden. NRF2-activering speelt een rol in het fase II-ontgiftingssysteem. Fase II-ontgifting neemt lipofiele of vetoplosbare vrije radicalen en zet deze om in hydrofiele of wateroplosbare stoffen voor uitscheiding, terwijl uitzonderlijk reactieve metabolieten en chemicaliën als gevolg daarvan worden geïnactiveerd van fase I.
NRF2-activering vermindert de algehele oxidatie en ontsteking van het menselijk lichaam door een hormooneffect. Om NRF2 te activeren, moet een ontstekingsreactie als gevolg van oxidatie optreden, zodat de cellen een adaptieve respons kunnen produceren en antioxidanten zoals glutathion kunnen aanmaken. Om het principe van Nrf2 te doorbreken, activeert oxidatieve stress in wezen NRF2, dat vervolgens een antioxidantreactie in het menselijk lichaam activeert. NRF2 functioneert om redox-signalering in evenwicht te brengen, of het evenwicht van oxidant- en antioxidantniveaus in de cel.
Een geweldige illustratie van hoe dit proces functioneert, kan worden gedemonstreerd met oefening. Bij elke training past de spier zich aan zodat deze geschikt is voor een andere trainingssessie. Als NRF2 onder- of overmatig tot expressie komt als gevolg van chronische infecties of verhoogde blootstelling aan toxines, wat kan worden waargenomen bij patiënten met het chronisch ontstekingsreactiesyndroom of CIRS, kunnen de gezondheidsproblemen verslechteren na NRF2-activering. Bovenal, als DJ-1 over-geoxideerd raakt, zal NRF2-activering te snel eindigen.
Effecten van NRF2-activering
NRF2-activering komt in hoge mate tot uiting in de longen, lever en nieren. Nucleaire erytroïde 2-gerelateerde factor 2 of NRF2 werkt meestal door het tegengaan van verhoogde niveaus van oxidatie in het menselijk lichaam die kunnen leiden tot oxidatieve stress. Nrf2-activering kan een aantal gezondheidsproblemen helpen behandelen, maar over-activering van Nrf2 kan verschillende problemen verergeren, die hieronder worden aangetoond.
Periodieke activering van Nrf2 kan helpen:
Veroudering (dwz levensduur)
Auto-immuniteit en algemene ontsteking (dwz artritis, autisme)
Kanker en Chemoprotection (ie EMF-blootstelling)
Depressie en angst (ie PTSS)
Blootstelling aan geneesmiddelen (alcohol, NSAID's)
Kanker (dwz hersenen, borst, hoofd, nek pancreas, prostaat, lever, schildklier)
Chronisch inflammatoir reactie-syndroom (CIRS)
Harttransplantatie (terwijl open NRF2 mogelijk slecht is, NRF2 kan helpen bij reparatie)
Hepatitis C
Nefritis (ernstige gevallen)
Vitiligo
Bovendien kan NRF2 helpen om specifieke voedingssupplementen, medicijnen en medicijnen te laten werken. Veel natuurlijke supplementen kunnen ook helpen om NRF2 te activeren. Door middel van huidige onderzoeken hebben onderzoekers aangetoond dat een groot aantal verbindingen waarvan ooit werd aangenomen dat ze antioxidanten waren, echt pro-oxidanten waren. Dat komt omdat ze bijna allemaal NRF2 nodig hebben om te functioneren, zelfs supplementen zoals curcumine en visolie. Cacao bleek bijvoorbeeld antioxiderende effecten te genereren bij muizen die het NRF2-gen bezitten.
Manieren om NRF2 te activeren
In het geval van neurodegeneratieve ziekten zoals de ziekte van Alzheimer, de ziekte van Parkinson, een beroerte of zelfs auto-immuunziekten, is het waarschijnlijk het beste om Nrf2 opgereguleerd te hebben, maar op een hormonale manier. Menging NRF2-activatoren kunnen ook een additief of synergistisch effect hebben, omdat het af en toe dosisafhankelijk kan zijn. De beste manieren om de Nrf2-expressie te vergroten, worden hieronder weergegeven:
Acupunctuur (dit is een alternatieve methode, laseracupunctuur kan ook worden gebruikt)
Vastend
Cannabidiol (CBD)
Lion's Mane + Melatonin
Alfa-liponzuur + DIM
Alsem
PPAR-gamma-activering
De volgende uitgebreide lijst met meer dan 350 andere manieren om Nrf2 te activeren via een dieet, levensstijl en apparaten, probiotica, supplementen, kruiden en oliën, hormonen en neurotransmitters, medicijnen / medicijnen en chemicaliën, pathways / transcriptiefactoren, en andere manieren, is alleen een korte handleiding over wat Nrf2 kan veroorzaken. Kortheidshalve hebben we in dit artikel andere 500-producten, voedingssupplementen en verbindingen die helpen om Nrf2 te activeren, weggelaten. De volgende zijn hieronder vermeld:
Dieet:
Acai bessen
Alcohol (Rode wijn is beter, vooral als er een kurk in zit, omdat protocatechuic aldehyde uit kurken ook NRF2 kan activeren.) Over het algemeen wordt alcohol niet aanbevolen, hoewel een acute inname NRF2 verhoogt. Chronische inname kan NRF2 verminderen.
Algen (kelp)
Appels
Zwarte thee
Paranoten
Broccoli Sprouts (en andere isothiocyanaten, sulforafaan en kruisbloemige groenten zoals paksoi met D3T)
Bosbessen (0.6-10 g / dag)
Wortelen (falcarinone)
Cayenne peper (Capsaïcine)
Selderij (butylftalide)
Chaga (Betulin)
Kamille thee
Chia
Chinese aardappel
Chokeberries (Aronia)
Chocolade (donker of cacao)
Kaneel
Koffie (zoals chlorogeenzuur, Cafestol en Kahweol)
Cordyceps
Vis (en schaaldieren)
Lijnzaad
Knoflook
Ghee (mogelijk)
Ginger (en Cardamonin)
Goji bessen
Grapefruit (Naringenin - 50 mg / kg / d naringenin)
Grapes
Groene thee
Guava
Hart van de Palm
Hijiki / Wakame
honingraat
kiwi
Peulvruchten
Manen van de leeuw
Mahuwa
Mango's (Mangiferin)
Mangisboom
Melk (geit, koe - via regulatie van het microbioom)
moerbeien
Olijfolie (afvallen - hydroxytyrosol en Oleanolic Acid)
Omega 6 vetzuren (Lipoxin A4)
Osange Oranges (Morin)
Oesterpaddestoelen
Papaja
Pinda's
Pigeon Peas
Granaatappel (Punicalagin, Ellagic Acid)
Propolis (Pinocembrin)
Paarse zoete aardappelen
Rambutan (Geraniin)
Uien
Reishi
Rhodiola Rosea (Salidroside)
Rice Bran (cycloartenyl ferulate)
Riceberry
Rooibos thee
Rozemarijn
Salie
saffloer
Sesamolie
Soja (en isoflavonen, Daidzein, Genistein)
squash
Aardbeien
Tartary Boekweit
Tijm
tomaten
Tonkabonen
Kurkuma
Wasabi
Watermelon
Lifestyle en apparaten:
Acupunctuur en elektro-acupunctuur (via collageencascade op ECM)
Oefening (Acute oefening zoals HIST of HIIT lijkt gunstiger te zijn voor het induceren van NRF2, terwijl langere inspanning NRF2 niet induceert, maar wel het glutathiongehalte verhoogt)
Vetarm dieet (dieet)
Hoge temperatuur (sauna)
Inhalatie van waterstof en waterstof Water
Hyperbare zuurstoftherapie
Infraroodtherapie (zoals Joovv)
Intraveneuze vitamine C
Ketogeen dieet
Ozon
Roken (niet aanbevolen - acuut roken verhoogt NRF2, chronisch roken vermindert NRF2. Als u ervoor kiest te roken, kan Holy Basil beschermen tegen downregulatie van NRF2)
Zon (UVB en infrarood)
probiotica:
Bacillus subtilis (fmbJ)
Clostridium butyricum (MIYAIRI 588)
Lactobacillus brevis
Lactobacillus casei (SC4 en 114001)
Lactobacillus collinoides
Lactobacillus gasseri (OLL2809, L13-Ia en SBT2055)
Lactobacillus helveticus (NS8)
Lactobacillus paracasei (NTU 101)
Lactobacillus plantarum (C88, CAI6, FC225, SC4)
Lactobacillus rhamnosus (GG)
Supplementen, kruiden en oliën:
Acetyl-L-Carnitine (ALCAR) en Carnitine
Allicine
Alfa-lipoïnezuur
amentoflavon
Andrographis paniculata
agmatine
Apigenin
Arginine
Artisjok (Cyanropicrin)
Ashwaganda
Astragalus
Bacopa
Biefstuk (Isogemaketone)
berberine
Beta-caryofylleen
Bidens Pilosa
Black Cumin Seed Oil (Thymoquinone)
Boswellia
Butein
butyraat
Cannabidiol (CBD)
Carotenioïden (zoals beta-caroteen [synergie met lycopeen – 2 - 15 mg/d lycopeen], fucoxanthine, zeaxanthine, astaxanthine en luteïne)
Chitrak
Chlorella
Chlorophyl
Chrysanthemum zawadskii
cinnamomea
Gemeenschappelijke zonnedauw
Koper
Coptis
CoQ10
Curcumine
Damiana
Dan Shen / Red Sage (Miltirone)
DIM
dioscine
Dong Ling Cao
Dong Quai (vrouwelijke ginseng)
Ecklonia Cava
EGCG
Elecampane / Inula
Eucommia schors
Ferulinezuur
fisetin
Visolie (DHA / EPA - 3 1 g / d visolie met 1098 mg EPA en 549 mg DHA)
Galangal
Gastrodin (Tian Ma)
Gentiana
Geranium
Ginkgo Biloba (Ginkgolide B)
zeekraal
Gotu Kola
Grape Seed Extract
Harige Agrimony
Haritaki (Triphala)
Hagedoorn
helichrysum
Henna (Juglone)
Hibiscus
Higenamine
Holy Basil / Tulsi (Ursolic Acid)
Hop
Horny Goat Weed (Icariin / Icariside)
Indigo Naturalis
IJzer (niet aanbevolen tenzij essentieel)
I3C
Job's Tears
Moringa Oleifera (zoals Kaempferol)
Inchinkoto (combinatie van Zhi Zi en Wormwood)
Kudzu Root
Licorice Root
Lindera Root
Luteoline (hoge doses voor activering, lagere doses remmen NRF2 bij kanker wel)
Magnolia
Manjistha
Maximowiczianum (Acerogenin A)
Mexicaanse Arnica
Milk Thistle
MitoQ
Mu Xiang
Mucuna Pruriens
Nicotinamide en NAD +
Panax Ginseng
Passiebloem (zoals Chrysin, maar chyrisine kan ook NRF2 verminderen via ontregeling van PI3K / Akt-signalering)
Resveratrol (hoofdzakelijk Piceid en andere fyto-oestrogenen, duizendknoop)
Rose Hips
Rozenhout
Rutine
Sappanwood
Sarsaparilla
Saururus chinensis
SC-E1 (Gips, jasmijn, zoethout, Kudzu en ballonbloem)
Schisandra
Self Heal (prunella)
Skullcap (Baicalin en Wogonin)
Sheep Sorrel
Si Wu Tang
Sideritis
Nardus (Aralia)
Spirulina
St. John's Wort
sulforafaan
Sutherlandia
Tao Hong Si Wu
Taurine
Thunder God Vine (Triptolide)
Tocoferolen (zoals vitamine E of Linalool)
Tribulus R
Tu Si Zi
TUDCA
Vitamine A (hoewel andere retinoïden NRF2 remmen)
Vitamine C (alleen hoge dosis, lage dosis remt NRF2)
Vitex / Chaste Tree
Witte pioen (Paeoniflorin van Paeonia lactiflora)
Alsem (Hispidulin en Artemisinin)
Xiao Yao Wan (Free and Easy Wanderer)
Yerba Santa (Eriodictyol)
Yuan Zhi (Tenuigenin)
Zi Cao (vermindert NRF2 bij kanker)
zink
Ziziphus Jujube
Hormonen en neurotransmitters:
adiponectin
Adropin
Oestrogeen (maar kan NRF2 verminderen in borstweefsel)
Melatonine
Progesteron
Quinolinezuur (in beschermende reactie om excitotoxiciteit te voorkomen)
Serotonine
Schildklierhormonen zoals T3 (kan NRF2 verhogen in gezonde cellen, maar verminderen bij kanker)
Vitamine D
Geneesmiddelen / medicijnen en chemicaliën:
Paracetamol
Acetazolamide
Amlodipine
auranofin
Bardoxolone methyl (BARD)
benznidazole
BHA
CDDO-imidazolide
Ceftriaxon (en bètalactamantibiotica)
Cialis
Dexamethason
Diprivan (Propofol)
eriodictyol
Exendine-4
Ezetimibe
Fluoride
fumaraat
HNE (geoxideerd)
idazoxan
Anorganisch arsenicum en natriumarseniet
JQ1 (kan ook NRF2 remmen, onbekend)
Letairis
melphalan
methazolamide
Methyleenblauw
Nifedipine
NSAID's
oltipraz
PPI's (zoals Omeprazol en Lansoprazol)
Protandim - geweldige resultaten in vivo, maar zwak / niet-bestaand bij het activeren van NRF2 bij mensen
Probucol
Rapamycine
Reserpine
rutenium
sitaxentan
Statines (zoals Lipitor en Simvastatin)
Tamoxifen
Tang Luo Ning
tBHQ
Tecfidera (dimethylfumaraat)
THC (niet zo sterk als CBD)
Theofylline
umbelliferon
Ursodeoxycholzuur (UDCA)
Verapamil
Viagra
4-Acetoxyphenol
Pathways / transcriptiefactoren:
7 nAChR-activering
AMPK
bilirubine
CDK20
CKIP-1
CYP2E1
EAATs
Gankyrin
kabouter
GJA1
H-ferritine ferroxidase
HDAC-remmers (zoals valproïnezuur en TSA, maar kunnen NRF2 instabiliteit veroorzaken)
Heat Shock Proteins
IL-17
IL-22
Klotho
let-7 (haalt mBach1 RNA naar beneden)
MAPK
Michael acceptors (meest)
miR-141
miR-153
miR-155 (haalt ook mBach1 RNA naar beneden)
miR-7 (in hersenen, helpt bij kanker en schizofrenie)
Notch1
Oxidatieve stress (zoals ROS, RNS, H2O2) en elektrofielen
PGC-1?
PKC-delta
PPAR-gamma (synergetische effecten)
Sigma-1-receptorremming
SIRT1 (verhoogt NRF2 in de hersenen en longen, maar kan dit in het algemeen verminderen)
SIRT2
SIRT6 (in de lever en hersenen)
SRXN1
TrxR1-remming (verzwakking of uitputting ook)
Zink protoporfyrine
4-HHE
Andere:
Ankaflavin
Asbest
Avicins
Bacillus amyloliquefaciens (gebruikt in de landbouw)
Koolmonoxide
Daphnetin
Glutathiondepletie (uitputting van 80%-90% mogelijk)
Gymnaster koraiensis
Hepatitis C
Herpes (HSV)
Indische es
Indigowoad Root
Isosalipurposide
Isorhamentin
Monascin
Omaveloxolone (sterk, ook bekend als RTA-408)
PDTC
Selenium-deficiëntie (seleniumdeficiëntie kan NRF2 verhogen)
Siberische Lariks
Sophoraflavanone G
Tadehagi triquetrum
Toona sinensis (7-DGD)
Trompet bloem
63171 en 63179 (sterk)
De nucleaire erytroïde 2-gerelateerde factor 2-signaleringsroute, het best bekend onder het acroniem Nrf2, is een transcriptiefactor die de belangrijkste rol speelt bij het reguleren van de beschermende antioxidantiemechanismen van het menselijk lichaam, in het bijzonder om oxidatieve stress onder controle te houden. Hoewel verhoogde niveaus van oxidatieve stress Nrf2 kunnen activeren, worden de effecten enorm verbeterd door de aanwezigheid van specifieke verbindingen. Bepaalde voedingsmiddelen en supplementen helpen Nrf2 in het menselijk lichaam te activeren, inclusief de isothiocyanaat sulforafaan van broccoli spruiten. Dr. Alex Jimenez DC, CCST Insight
Sulforafaan en de effecten ervan op kanker, mortaliteit, veroudering, hersenen en gedrag, hartaandoeningen en meer
Isothiocyanaten zijn enkele van de belangrijkste plantaardige stoffen die u in uw dieet kunt krijgen. In deze video maak ik de meest uitgebreide case voor hen die ooit is gemaakt. Korte aandachtsspanne? Ga naar uw favoriete onderwerp door op een van de onderstaande tijdspunten te klikken. Volledige tijdlijn hieronder.
Belangrijkste secties:
00: 01: 14 - Kanker en sterfte
00: 19: 04 - Veroudering
00: 26: 30 - Hersenen en gedrag
00: 38: 06 - Laatste overzicht
00: 40: 27 - Dosis
Volledige tijdlijn:
00: 00: 34 - Introductie van sulforafaan, een belangrijk aandachtspunt van de video.
00: 01: 14 - Kruisbloemige groenteconsumptie en verlagingen van mortaliteit door alle oorzaken.
00: 02: 12 - Risico op prostaatkanker.
00: 02: 23 - Blaaskankerrisico.
00: 02: 34 - Longkanker bij rokers risico.
00: 02: 48 - Risico op borstkanker.
00: 03: 13 - Hypothetisch: wat als je al kanker hebt? (Interventie)
00: 03: 35 - Aannemelijk mechanisme dat de associatieve gegevens over kanker en mortaliteit stuurt.
00: 04: 38 - Sulforafaan en kanker.
00: 05: 32 - Dierlijk bewijs dat een sterk effect van extract van broccolispruit toont op de ontwikkeling van blaastumoren bij ratten.
00: 06: 06 - Effect van directe suppletie van sulforafaan bij prostaatkankerpatiënten.
00: 07: 09 - Bioaccumulatie van isothiocyanaatmetabolieten in daadwerkelijk borstweefsel.
00: 08: 32 - Remming van stamcellen van borstkanker.
00: 08: 53 - Geschiedenisles: brassica's werden vastgesteld met gezondheidseigenschappen zelfs in het oude Rome.
00: 09: 16 - het vermogen van Sulforaphane om de kankerverwekkende uitscheiding te verbeteren (benzeen, acroleïne).
00: 09: 51 - NRF2 als een genetische switch via antioxidantresponselementen.
00: 10: 10 - Hoe NRF2-activering de carcinogene excretie via glutathion-S-conjugaten verbetert.
00: 10: 34 - Spruitjes verhogen glutathion-S-transferase en verminderen DNA-schade.
00: 11: 20 - Broccolispruitendrank verhoogt de benzeenuitscheiding met 61%.
00: 13: 31 - Broccoli-spruithomogenaat verhoogt antioxiderende enzymen in de bovenste luchtwegen.
00: 15: 45 - Kruisbloemige groenteconsumptie en sterfte aan hart- en vaatziekten.
00: 16: 55 - Broccolispruitpoeder verbetert de bloedlipiden en het algemene risico op hartziekten bij type 2 diabetici.
00: 19: 04 - Begin van het verouderingsgedeelte.
00: 19: 21 - Met Sulforaphane verrijkte voeding verlengt de levensduur van kevers van 15 tot 30% (onder bepaalde omstandigheden).
00: 20: 34 - Belang van lage ontsteking voor een lang leven.
00: 22: 05 - Kruisbloemige groenten en broccoli-kiempoeder lijken een breed scala aan ontstekingsmarkers bij mensen te verminderen.
00: 24: 14 - Muisstudies suggereren dat sulforaphane de adaptieve immuunfunctie op oudere leeftijd zou kunnen verbeteren.
00: 25: 18 - Sulforafaan verbeterde de haargroei in een muismodel van kalend. Afbeelding bij 00: 26: 10.
00: 26: 30 - Begin van de sectie hersenen en gedrag.
00: 27: 18 - Effect van extract van broccolispruiten op autisme.
00: 27: 48 - Effect van glucoraphanine op schizofrenie.
00: 28: 17 - Begin van depressie discussie (plausibel mechanisme en studies).
00: 31: 21 - Muisstudie met behulp van 10 verschillende modellen van stress-geïnduceerde depressie laten sulforafaan zien, even effectief als fluoxetine (prozac).
00: 32: 00 - Onderzoek toont aan dat directe inname van glucoraphanine bij muizen even effectief is bij het voorkomen van depressies als gevolg van een sociaal nederlaagstressmodel.
00: 33: 01 - Begin van de sectie neurodegeneratie.
00: 33: 30 - Sulforafane en de ziekte van Alzheimer.
00: 33: 44 - Sulforafane en de ziekte van Parkinson.
00: 33: 51 - De ziekte van Sulforaphane en Hungtington.
00: 34: 43 - Begin van een traumatisch gedeelte over hersenletsel.
00: 35: 01 - Sulforafaan wordt geïnjecteerd onmiddellijk nadat TBI het geheugen verbetert (muisstudie).
00: 35: 55 - Sulforafaan en neuronale plasticiteit.
00: 36: 32 - Sulforafaan verbetert het leren in het model van type II diabetes bij muizen.
00: 37: 19 - Sulforafaan en duchenne spierdystrofie.
00: 37: 44 - Myostatinremming in spieratlanticellen (in vitro).
00: 38: 06 - Late video recapitulatie: sterfte en kanker, DNA-schade, oxidatieve stress en ontsteking, uitscheiding van benzeen, cardiovasculaire aandoeningen, type II diabetes, effecten op de hersenen (depressie, autisme, schizofrenie, neurodegeneratie), NRF2-route.
00: 40: 27 - Gedachten over het uitzoeken van een dosis broccoli of sulforafaan.
00: 41: 01 - Anekdotes over ontspruiten thuis.
00: 43: 14 - Op kooktemperaturen en sulforafaanactiviteit.
00: 43: 45 - Darmbacterieomzetting van sulforafaan uit glucoraphanine.
00: 44: 24 - Supplementen werken beter in combinatie met actieve myrosinase van groenten.
00: 44: 56 - Koken technieken en kruisbloemige groenten.
00: 46: 06 - Isothiocyanaten als goitrogens.
Volgens veel huidige onderzoeken is de nucleaire erytroïde 2-gerelateerde factor 2-signaleringsroute, beter bekend als Nrf2, een fundamentele transcriptiefactor die de beschermende antioxidantmechanismen van de cellen activeert om het menselijk lichaam te ontgiften van zowel externe als interne factoren en om verhoogde niveaus van oxidatieve stress. De reikwijdte van onze informatie is beperkt tot chiropractie en problemen met de wervelkolom. Om het onderwerp te bespreken, kunt u het aan Dr. Jimenez vragen of contact met ons opnemen via 915-850-0900 .
Curator van Dr. Alex Jimenez
Bijkomend onderwerpbespreking: Acute rugpijn
Rugpijn is een van de meest voorkomende oorzaken van arbeidsongeschiktheid en gemiste werkdagen wereldwijd. Rugpijn is de tweede meest voorkomende reden voor doktersbezoeken, alleen overtroffen door bovenste luchtweginfecties. Ongeveer 80 procent van de bevolking zal gedurende zijn leven minstens één keer rugpijn ervaren. De wervelkolom is een complexe structuur die bestaat uit botten, gewrichten, ligamenten en spieren, naast andere zachte weefsels. Letsel en / of verergerde omstandigheden, zoals herniated discs, kan uiteindelijk leiden tot symptomen van rugpijn. Sportblessures of auto-ongelukken zijn vaak de meest voorkomende oorzaak van rugpijn, maar soms kunnen de eenvoudigste bewegingen pijnlijke gevolgen hebben. Gelukkig kunnen alternatieve behandelingsopties, zoals chiropractische zorg, rugpijn helpen verlichten door het gebruik van spinale aanpassingen en handmatige manipulaties, waardoor uiteindelijk de pijnverlichting wordt verbeterd. �
Oxidatieve stress is een belangrijke oorzaak van de ontwikkeling van een verscheidenheid aan gezondheidsproblemen, waaronder kanker, hartaandoeningen, diabetes, versnelde veroudering en neurodegeneratie. Voedingsmiddelen, kruiden en supplementen die rijk zijn aan antioxidanten kunnen worden gebruikt om het menselijk lichaam te beschermen tegen hoge niveaus van oxidatieve stress. Recente onderzoeken hebben aangetoond dat de Nrf2-genroute kan helpen de effecten van antioxidanten te versterken. De voordelen van Nrf2 worden hieronder beschreven.
Beschermt het lichaam tegen gifstoffen
NRF2 is een intrinsieke stof die de cellen kan beschermen tegen schadelijke, interne en externe verbindingen. NRF2 kan helpen de reactie van het menselijk lichaam op medicijnen/medicijnen en toxines te verrijken, waardoor de productie van eiwitten wordt verbeterd die helpen bij het elimineren van verbindingen uit de cel, bekend als multidrug-resistentie-geassocieerde eiwitten of MRP's. NRF2 wordt bijvoorbeeld geactiveerd bij sigarettenrook inademen om de longen te laten ontgiften.
Bovendien is het essentieel voor de longen om zichzelf te beschermen tegen allergenen, virale ziekten, bacteriële endotoxinen, hyperoxie en verschillende milieuverontreinigende stoffen. De constante trigger van Nrf2 kan echter de niveaus van een stof die bekend staat als glutathion in het menselijk lichaam verlagen. NRF2 kan ook de lever beschermen tegen toxiciteit en het kan de lever beschermen tegen arseenhepatotoxiciteit. Bovendien beschermt NRF2 de lever en hersenen tegen alcoholgebruik. Nrf2 kan bijvoorbeeld beschermen tegen acetaminophen-toxiciteit.
Bestrijdt ontstekingen en oxidatieve stress
NRF2-activering kan ontstekingen helpen bestrijden door inflammatoire cytokines, zoals die aanwezig zijn bij psoriasis, te verminderen. NRF2 kan ook ontstekingen verminderen die gepaard gaan met een verscheidenheid aan gezondheidsproblemen, zoals artritis en fibrose van de lever, nieren en longen. NRF2 kan ook helpen bij het beheersen van allergieën door Th1/Th17-cytokinen te verlagen en TH2-cytokinen te verhogen. Dit kan gunstig zijn voor aandoeningen zoals astma.
NRF2 beschermt bovendien tegen celbeschadiging door blauw licht en tegen UVA/UVB in zonlicht. Nrf2-tekortkomingen kunnen het een stuk gemakkelijker maken om door de zon te verbranden. Een reden hiervoor is dat NRF2 collageen kan reguleren als reactie op UV-straling. Advanced Glycation End-Products, of AGE's, dragen bij aan de ontwikkeling van veel gezondheidsproblemen, waaronder diabetes en neurodegeneratieve ziekten. NRF2 kan de oxidatieve stress van AGE's in het lichaam verminderen. NRF2 kan het menselijk lichaam ook beschermen tegen hogere niveaus van op hitte gebaseerde stress.
Verbetert mitochondriën en trainingsprestaties
NRF2 is een mitochondriale booster. NRF2-activering draagt bij aan een stijging van de ATP-energie voor mitochondriën, naast een verbeterd gebruik van zuurstof, of citraat, en vet. Zonder NRF2 zouden mitochondriën gewoon kunnen functioneren met suiker of glucose in plaats van vet. NRF2 is ook essentieel voor de ontwikkeling van mitochondriën via een proces dat bekend staat als biogenese. NRF2-activering is van vitaal belang om te profiteren van de voordelen van lichaamsbeweging.
Vanwege de activiteit van Nrf2 verhoogt lichaamsbeweging de mitochondriale functie, waar dit resultaat kan worden versterkt met CoQ10, Cordyceps en caloriebeperking. Matige lichaamsbeweging of acute lichaamsbeweging induceert mitochondriale biogenese en een verhoogde synthese van superoxide dismutase, of SOD, en heem-oxygenase-1, of HO-1, door NRF2-activering. Alfa-liponzuur, of ALA, en Dan Shen kunnen de door NRF2 gemedieerde mitochondriale biogenese stimuleren. Bovendien kan NRF2 ook de inspanningstolerantie verbeteren wanneer NRF2-verwijdering training schadelijk maakt.
Beschermt tegen hypoxie
NRF2 helpt ook het menselijk lichaam te beschermen tegen cellulair zuurstofverlies/-uitputting, een gezondheidsprobleem dat hypoxie wordt genoemd. Personen met CIRS hebben verminderde zuurstofniveaus omdat hun NRF2 wordt belemmerd, wat resulteert in verminderde niveaus van zowel VEGF, HIF1 als HO-1. Gewoonlijk wordt miR-101, dat nodig is voor de aanmaak van stamcellen, bij gezonde personen met hypoxie tot overexpressie gebracht en verhoogt het de hoeveelheden NRF2/HO-1 en VEGF/eNOS, waardoor hersenbeschadiging wordt voorkomen, maar dat lijkt niet te gebeuren in CIRS.
Hypoxie, gekenmerkt door lage HIF1, in CIRS kan ook resulteren in een lekkende bloed-hersenbarrière als gevolg van een NRF2-onbalans. Salidroside, gelegen in de Rhodiola, werkt op NRF2-activering en helpt bij hypoxie door de niveaus van VEGF en HIF1 in het menselijk lichaam te verhogen. NRF2 kan uiteindelijk ook beschermen tegen lactaatophoping in het hart. NRF2-activering kan ook door hypoxie veroorzaakte hoogteziekte of AMS stoppen.
Vertraagt veroudering
Verschillende verbindingen die in grote hoeveelheden dodelijk kunnen zijn, kunnen de levensduur in vrij kleine hoeveelheden verlengen als gevolg van xenohormese via NRF2, PPAR-gamma en FOXO. Een zeer kleine hoeveelheid toxines verhoogt het vermogen van de cel om beter uitgerust te worden voor de volgende keer dat het wordt uitgedaagd met een toxine, maar dit is geen goedkeuring om giftige chemicaliën te consumeren.
Een goede illustratie van dit proces is met caloriebeperking. NRF2 kan de levensduur van cellen verbeteren door hun niveaus van mitochondriën en antioxidanten te verhogen en het vermogen van de cellen om te sterven te verlagen. NRF2 neemt af met het ouder worden omdat NRF2 voorkomt dat stamcellen afsterven en hen helpt te regenereren. NRF2 speelt een rol bij het verbeteren van wondgenezing.
Versterkt het vaatstelsel
Correct uitgevoerd met de productie van sulforafaan, kan NRF2-activering beschermen tegen hartziekten zoals hoge bloeddruk of hypertensie en verharding van de slagaders of atherosclerose. NRF2 kan de ontspannende activiteit van Acetylcholine of ACh op het vasculaire systeem versterken en tegelijkertijd door cholesterol veroorzaakte stress verminderen. Nrf2-activering kan het hart versterken, maar overgeactiveerde Nrf2 kan de kans op hart- en vaatziekten verhogen.
Statines kunnen hart- en vaatziekten voorkomen of ertoe leiden. NRF2 speelt ook een belangrijke rol bij het balanceren van ijzer en calcium, wat het menselijk lichaam kan beschermen tegen verhoogde ijzerspiegels. Sirtuin 2, of SIRT2, kan bijvoorbeeld de ijzerhomeostase in cellen reguleren door activering van NRF2, waarvan wordt aangenomen dat het nodig is voor een gezond ijzergehalte. NRF2 kan ook helpen bij sikkelcelziekte of SCD. NRF2-disfunctie kan een reden zijn voor endotoxemie, zoals bij dysbiose of door lectines geïnduceerde hypertensie. Nrf2 kan het menselijk lichaam ook beschermen tegen door amfetamine veroorzaakte schade aan het vaatstelsel.
Bestrijdt neuro-inflammatie
NRF2 kan beschermen tegen en helpen bij ontsteking van de hersenen, gewoonlijk neuro-inflammatie genoemd. Bovendien kan NRF2 helpen bij een assortiment van aandoeningen van het centrale zenuwstelsel of het CZS, waaronder:
Ziekte van Alzheimer (AD) - vermindert amyloïde bètastress op mitochondriën
Amyotrofische laterale sclerose (ALS)
Ziekte van Huntington (HD)
Multiple sclerose (MS)
Zenuwregeneratie
Ziekte van Parkinson (PD) – beschermt dopamine
Ruggenmergletsel (SCI)
Beroerte (ischemisch en hemorragisch) - helpt hypoxie
Traumatisch Hersenletsel
NRF2 heeft een afname van neuro-inflammatie onthuld bij tieners met autismespectrumstoornissen of ASS. Idebenone gaat goed samen met NRF2-activators in tegenstelling tot neuro-inflammatie. NRF2 kan ook de Blood Brain Barrier, of BBB, verbeteren. NRF2-activering met carnosinezuur verkregen uit rozemarijn en salie kan bijvoorbeeld de BBB passeren en neurogenese veroorzaken. Van NRF2 is ook aangetoond dat het de Brain Derived Neurotrophic Factor of BDNF verhoogt.
NRF2 moduleert ook het vermogen van sommige voedingssupplementen om zenuwgroeifactor of NGF te veroorzaken, omdat het ook kan helpen bij hersenmist en glutamaat-geïnduceerde problemen door N-methyl-D-aspartaat of NMDA-receptoren te moduleren. Het kan ook de oxidatieve stress van chinolinezuur verlagen, ook wel QUIN genoemd. NRF2-activering kan beschermen tegen aanvallen en hoge doses kunnen de rand van een aanval verminderen. Bij regelmatige stimulatiedoses kan NRF2 de cognitieve vermogens verbeteren na een aanval door extracellulair glutamaat in de hersenen te verlagen en door het vermogen om cysteïne uit glutamaat en glutathion te halen.
Verlicht depressie
Bij depressie is het normaal om ontstekingen in de hersenen op te merken, vooral van de prefrontale cortex en de hippocampus, evenals verminderde BDNF. In sommige versies van depressie kan NRF2 depressieve symptomen verbeteren door de ontsteking in de hersenen te verlagen en de BDNF-spiegels te verhogen. Het vermogen van Agmatine om depressie te verminderen door noradrenaline, dopamine, serotonine en BDNF in de hippocampus te verhogen, hangt af van NRF2-activering.
Bevat eigenschappen tegen kanker
NRF2 is evenzeer een tumorsuppressor als een tumorpromotor als het niet dienovereenkomstig wordt behandeld. NRF2 kan beschermen tegen kanker veroorzaakt door vrije radicalen en oxidatieve stress, maar overexpressie van NRF2 kan ook in kankercellen worden gevonden. Intense activering van NRF2 kan helpen bij een verscheidenheid aan kankers. Zo kan het supplement Protandim huidkanker verminderen door NRF2-activering.
Verzacht de pijn
Golfoorlogziekte, of GWI, een opmerkelijke ziekte die Golfoorlogveteranen treft, is een verzameling onverklaarbare, chronische symptomen die vermoeidheid, hoofdpijn, gewrichtspijn, indigestie, slapeloosheid, duizeligheid, ademhalingsproblemen en geheugenproblemen kunnen omvatten. NRF2 kan de symptomen van GWI verbeteren door de hippocampus en algemene ontsteking te verminderen, naast het verminderen van pijn. NRF2 kan bovendien helpen bij pijn door lichamelijk zenuwletsel en zenuwbeschadiging door diabetische neuropathie verbeteren.
Verbetert diabetes
Hoge glucosespiegels, het best aangeduid als hyperglykemie, veroorzaken oxidatieve schade aan de cellen als gevolg van de verstoring van de mitochondriale functie. NRF2-activering kan het menselijk lichaam beschermen tegen de schade van hyperglykemie aan de cel, waardoor celdood wordt voorkomen. NRF2-activering kan bovendien de bètacelfunctie van de pancreas beschermen, herstellen en verbeteren, terwijl de insulineresistentie wordt verminderd.
Beschermt zicht en gehoor
NRF2 kan beschermen tegen schade aan het oog door diabetische retinopathie. Het kan ook de vorming van cataracten voorkomen en fotoreceptoren beschermen in tegenstelling tot door licht veroorzaakte dood. NRF2 beschermt bovendien het oor, of cochlea, tegen stress en gehoorverlies.
Kan obesitas helpen
NRF2 kan helpen bij obesitas, voornamelijk vanwege het vermogen om variabelen te reguleren die werken op vetophoping in het menselijk lichaam. NRF2-activering met sulforafaan kan de remming van vetzuursynthese, of FAS, en ontkoppelingseiwitten, of UCP, verhogen, wat resulteert in minder vetophoping en meer bruin vet, dat wordt gekenmerkt als vet dat meer mitochondriën bevat.
Beschermt de darm
NRF2 helpt de darm te beschermen door de homeostase van het darmmicrobioom te beschermen. Lactobacillus-probiotica zullen bijvoorbeeld NRF2 activeren om de darm te beschermen tegen oxidatieve stress. NRF2 kan ook helpen bij het voorkomen van colitis ulcerosa of UC.
Beschermt geslachtsorganen
NRF2 kan de testikels beschermen en het aantal zaadcellen beschermen tegen schade bij mensen met diabetes. Het kan ook helpen bij erectiestoornissen of ED. Sommige libido-verhogende supplementen zoals Mucuna, Tribulus en Ashwaganda kunnen de seksuele functie verbeteren via NRF2-activering. Andere factoren die NRF2 stimuleren, zoals zonlicht of broccolispruiten, kunnen ook helpen het libido te verbeteren.
Reguleert botten en spieren
Oxidatieve stress kan leiden tot vermindering van de botdichtheid en sterkte, wat normaal is bij osteoporose. NRF2-activering zou het vermogen kunnen hebben om antioxidanten in botten te verbeteren en te beschermen tegen botveroudering. NRF2 kan ook spierverlies voorkomen en Duchenne spierdystrofie of DMD versterken.
Bevat antivirale eigenschappen
Last but not least kan NRF2-activering uiteindelijk helpen het menselijk lichaam te verdedigen tegen verschillende virussen. Bij patiënten met het dengue-virus waren de symptomen niet zo intens bij personen met hogere NRF2-waarden dan bij personen met minder NRF2-waarden. NRF2 kan ook mensen helpen die het humaan immunodeficiëntie-1-virus of hiv hebben. NRF2 kan beschermen tegen de oxidatieve stress van Adeno-Associated Virus, of AAV, en H. Pylori. Ten slotte kan Lindera Root het hepatitis C-virus onderdrukken met NRF2-activering.
Nrf2, of NF-E2-gerelateerde factor 2, is een transcriptiefactor die bij mensen wordt aangetroffen en die de expressie van een specifieke reeks antioxidant- en ontgiftende genen reguleert. Deze signaalroute wordt geactiveerd als gevolg van oxidatieve stress, omdat het talrijke antioxidanten en fase II-leverontgiftingsenzymen verbetert om de homeostase in het menselijk lichaam te herstellen. Mensen zijn aangepast om te functioneren in een staat van homeostase of evenwicht. Wanneer het lichaam wordt geconfronteerd met oxidatieve stress, wordt Nrf2 geactiveerd om oxidatie te reguleren en de stress die het veroorzaakt te beheersen. Nrf2 is essentieel om gezondheidsproblemen die verband houden met oxidatieve stress te voorkomen. Dr. Alex Jimenez DC, CCST Insight
Sulforafaan en de effecten ervan op kanker, mortaliteit, veroudering, hersenen en gedrag, hartaandoeningen en meer
Isothiocyanaten zijn enkele van de belangrijkste plantaardige stoffen die u in uw dieet kunt krijgen. In deze video maak ik de meest uitgebreide case voor hen die ooit is gemaakt. Korte aandachtsspanne? Ga naar uw favoriete onderwerp door op een van de onderstaande tijdspunten te klikken. Volledige tijdlijn hieronder.
Belangrijkste secties:
00: 01: 14 - Kanker en sterfte
00: 19: 04 - Veroudering
00: 26: 30 - Hersenen en gedrag
00: 38: 06 - Laatste overzicht
00: 40: 27 - Dosis
Volledige tijdlijn:
00: 00: 34 - Introductie van sulforafaan, een belangrijk aandachtspunt van de video.
00: 01: 14 - Kruisbloemige groenteconsumptie en verlagingen van mortaliteit door alle oorzaken.
00: 02: 12 - Risico op prostaatkanker.
00: 02: 23 - Blaaskankerrisico.
00: 02: 34 - Longkanker bij rokers risico.
00: 02: 48 - Risico op borstkanker.
00: 03: 13 - Hypothetisch: wat als je al kanker hebt? (Interventie)
00: 03: 35 - Aannemelijk mechanisme dat de associatieve gegevens over kanker en mortaliteit stuurt.
00: 04: 38 - Sulforafaan en kanker.
00: 05: 32 - Dierlijk bewijs dat een sterk effect van extract van broccolispruit toont op de ontwikkeling van blaastumoren bij ratten.
00: 06: 06 - Effect van directe suppletie van sulforafaan bij prostaatkankerpatiënten.
00: 07: 09 - Bioaccumulatie van isothiocyanaatmetabolieten in daadwerkelijk borstweefsel.
00: 08: 32 - Remming van stamcellen van borstkanker.
00: 08: 53 - Geschiedenisles: brassica's werden vastgesteld met gezondheidseigenschappen zelfs in het oude Rome.
00: 09: 16 - het vermogen van Sulforaphane om de kankerverwekkende uitscheiding te verbeteren (benzeen, acroleïne).
00: 09: 51 - NRF2 als een genetische switch via antioxidantresponselementen.
00: 10: 10 - Hoe NRF2-activering de carcinogene excretie via glutathion-S-conjugaten verbetert.
00: 10: 34 - Spruitjes verhogen glutathion-S-transferase en verminderen DNA-schade.
00: 11: 20 - Broccolispruitendrank verhoogt de benzeenuitscheiding met 61%.
00: 13: 31 - Broccoli-spruithomogenaat verhoogt antioxiderende enzymen in de bovenste luchtwegen.
00: 15: 45 - Kruisbloemige groenteconsumptie en sterfte aan hart- en vaatziekten.
00: 16: 55 - Broccolispruitpoeder verbetert de bloedlipiden en het algemene risico op hartziekten bij type 2 diabetici.
00: 19: 04 - Begin van het verouderingsgedeelte.
00: 19: 21 - Met Sulforaphane verrijkte voeding verlengt de levensduur van kevers van 15 tot 30% (onder bepaalde omstandigheden).
00: 20: 34 - Belang van lage ontsteking voor een lang leven.
00: 22: 05 - Kruisbloemige groenten en broccoli-kiempoeder lijken een breed scala aan ontstekingsmarkers bij mensen te verminderen.
00: 24: 14 - Muisstudies suggereren dat sulforaphane de adaptieve immuunfunctie op oudere leeftijd zou kunnen verbeteren.
00: 25: 18 - Sulforafaan verbeterde de haargroei in een muismodel van kalend. Afbeelding bij 00: 26: 10.
00: 26: 30 - Begin van de sectie hersenen en gedrag.
00: 27: 18 - Effect van extract van broccolispruiten op autisme.
00: 27: 48 - Effect van glucoraphanine op schizofrenie.
00: 28: 17 - Begin van depressie discussie (plausibel mechanisme en studies).
00: 31: 21 - Muisstudie met behulp van 10 verschillende modellen van stress-geïnduceerde depressie laten sulforafaan zien, even effectief als fluoxetine (prozac).
00: 32: 00 - Onderzoek toont aan dat directe inname van glucoraphanine bij muizen even effectief is bij het voorkomen van depressies als gevolg van een sociaal nederlaagstressmodel.
00: 33: 01 - Begin van de sectie neurodegeneratie.
00: 33: 30 - Sulforafane en de ziekte van Alzheimer.
00: 33: 44 - Sulforafane en de ziekte van Parkinson.
00: 33: 51 - De ziekte van Sulforaphane en Hungtington.
00: 34: 43 - Begin van een traumatisch gedeelte over hersenletsel.
00: 35: 01 - Sulforafaan wordt geïnjecteerd onmiddellijk nadat TBI het geheugen verbetert (muisstudie).
00: 35: 55 - Sulforafaan en neuronale plasticiteit.
00: 36: 32 - Sulforafaan verbetert het leren in het model van type II diabetes bij muizen.
00: 37: 19 - Sulforafaan en duchenne spierdystrofie.
00: 37: 44 - Myostatinremming in spieratlanticellen (in vitro).
00: 38: 06 - Late video recapitulatie: sterfte en kanker, DNA-schade, oxidatieve stress en ontsteking, uitscheiding van benzeen, cardiovasculaire aandoeningen, type II diabetes, effecten op de hersenen (depressie, autisme, schizofrenie, neurodegeneratie), NRF2-route.
00: 40: 27 - Gedachten over het uitzoeken van een dosis broccoli of sulforafaan.
00: 41: 01 - Anekdotes over ontspruiten thuis.
00: 43: 14 - Op kooktemperaturen en sulforafaanactiviteit.
00: 43: 45 - Darmbacterieomzetting van sulforafaan uit glucoraphanine.
00: 44: 24 - Supplementen werken beter in combinatie met actieve myrosinase van groenten.
00: 44: 56 - Koken technieken en kruisbloemige groenten.
00: 46: 06 - Isothiocyanaten als goitrogens.
Wanneer het menselijk lichaam wordt geconfronteerd met schadelijke interne en externe factoren zoals toxines, moeten de cellen snel hun antioxiderende eigenschappen activeren om oxidatieve stress tegen te gaan. Omdat is vastgesteld dat verhoogde niveaus van oxidatieve stress verschillende gezondheidsproblemen veroorzaken, is het belangrijk om Nrf2-activering te gebruiken om van de voordelen te profiteren. De reikwijdte van onze informatie is beperkt tot chiropractische en spinale gezondheidsproblemen. Als u het onderwerp wilt bespreken, kunt u het aan Dr. Jimenez vragen of contact met ons opnemen via�915-850-0900 .
Curator van Dr. Alex Jimenez
Bijkomend onderwerpbespreking: Acute rugpijn
Rugpijn is een van de meest voorkomende oorzaken van arbeidsongeschiktheid en gemiste werkdagen wereldwijd. Rugpijn is de tweede meest voorkomende reden voor doktersbezoeken, alleen overtroffen door bovenste luchtweginfecties. Ongeveer 80 procent van de bevolking zal gedurende zijn leven minstens één keer rugpijn ervaren. De wervelkolom is een complexe structuur die bestaat uit botten, gewrichten, ligamenten en spieren, naast andere zachte weefsels. Hierdoor kunnen verwondingen en / of verergerde omstandigheden, zoals herniated discs, kan uiteindelijk leiden tot symptomen van rugpijn. Sportblessures of auto-ongelukken zijn vaak de meest voorkomende oorzaak van rugpijn, maar soms kunnen de eenvoudigste bewegingen pijnlijke gevolgen hebben. Gelukkig kunnen alternatieve behandelingsopties, zoals chiropractische zorg, rugpijn helpen verlichten door het gebruik van spinale aanpassingen en handmatige manipulaties, waardoor uiteindelijk de pijnverlichting wordt verbeterd. �
sulforafaan is een fytochemische stof, een stof binnen de isothiocyanaatgroep van organozwavelverbindingen, aangetroffen in kruisbloemige groenten, zoals broccoli, kool, bloemkool en spruitjes. Het kan ook worden gevonden in paksoi, boerenkool, collards, mosterdgroenten en waterkers. Onderzoeksstudies hebben aangetoond dat sulforafaan verschillende vormen van kanker kan helpen voorkomen activeren van de productie van Nrf2, of nucleaire factor erytroïde 2-gerelateerde factor, een transcriptiefactor die beschermende antioxidantmechanismen reguleert die de reactie van de cel op oxidanten regelen. Het doel van het volgende artikel is om de functie van sulforafaan te beschrijven.
Abstract
Het KEAP1-NrF2-ARE antioxidantensysteem is een belangrijk middel waarmee cellen reageren op oxidatieve en xenobiotische spanningen. Sulforaphane (SFN), een elektrofiel isothiocyanaat afgeleid van kruisbloemige groenten, activeert de KEAP1-NrF2-ARE-route en is een moleculair belang geworden bij de behandeling van ziekten waarbij chronische oxidatieve stress een belangrijke etiologische rol speelt. We demonstreren hier dat de mitochondriën van gekweekte, humane retinale pigmentepitheelcellen (RPE-1) die met SFN worden behandeld, een hyperfusie ondergaan die onafhankelijk is van zowel Nrf2 als zijn cytoplasmatische remmer KEAP1. Mitochondriale fusie is naar verluidt cytoprotectief door het remmen van porievorming in mitochondria tijdens apoptose, en in overeenstemming hiermee laten we Nrf2-onafhankelijke cytoprotectie zien van met SFN behandelde cellen die zijn blootgesteld aan de apoptose-inductor, staurosporine. Mechanistisch compenseert SFN de rekrutering en / of retentie van de oplosbare splijtingsfactor Drp1 naar mitochondria en peroxisomen, maar heeft geen invloed op de algehele overvloed van Drp1. Deze gegevens tonen aan dat de gunstige eigenschappen van SFN verder reiken dan de activering van het KEAP1-Nrf2-ARE-systeem en verdere ondervraging rechtvaardigen gezien het huidige gebruik van dit middel in meerdere klinische onderzoeken.
Sulforaphane is een Nrf2-onafhankelijke remmer van mitochondriale splijting
Sulforafaan (SFN) is een isothiocyanaatverbinding afgeleid van het dieet dat het meest voorkomt uit kruisbloemige groenten [56]. Het wordt gegenereerd in planten als een xenobiotische reactie op predatie via vesiculaire afgifte van het hydrolytische enzym myrosinase uit beschadigde cellen; dit enzym zet glucosinolaten om in isothiocyanen [42]. In de afgelopen twee decennia is SFN uitgebreid gekarakteriseerd vanwege de gerapporteerde antikanker-, antioxidant- en antimicrobiële eigenschappen [57]. Veel van deze werkzaamheid is toegeschreven aan het vermogen van SFN om de KEAP1-Nrf2-antioxidant responselement (ARE) signaalroute te moduleren, hoewel aanvullende activiteiten van de verbinding zijn geïdentificeerd, waaronder de remming van histondeacetylase-activiteit en celcyclusprogressie [ 29]. Nrf2 is de meest belangrijke transcriptiefactor voor antioxidanten en onder omstandigheden van homeostase wordt de stabiliteit ervan onderdrukt door de werking van het cytoplasmische Cullin3KEAP1 ubiquitine-ligasecomplex [20]. In het bijzonder wordt Nrf2 gerekruteerd naar de Cullin3KEAP1-ligase door binding aan de dimere substraatadapter KEAP1 en wordt vervolgens gemodificeerd met polyUb-ketens die de transcriptiefactor voor proteasoom-gemedieerde afbraak richten. Deze constitutieve turnover beperkt de halfwaardetijd van Nrf2 in onbeklede cellen tot ~ 15 min [30], [33], [46], [55]. In reactie op talrijke soorten stress, met name oxidatieve stress, werkt KEAP1, een cysteïnerijk proteïne, als een redoxsensor, en oxidatieve modificatie van kritieke cysteïnes, met name C151, van KEAP1 dissocieert Nrf2-KEAP1 van CUL3 waardoor Nrf2-degradatie wordt voorkomen [ 8], [20], [55]. Met name SFN en mogelijk andere Nrf2-activatoren bootsen oxidatieve stress na door C151 van KEAP1 te wijzigen, bijvoorbeeld [21]. Stabilisatie van Nrf2 zorgt voor de translocatie naar de kern waar het de expressie induceert van een batterij van Phase II antioxidant- en detoxificatiegenen. Nrf2 bindt zich aan de antioxidantresponspromotorelementen (ARE) van zijn verwante doelwitgenen door heterodimerisatie met kleine Maf-eiwitten [19]. Dit systeem presenteert een dynamische en gevoelige reactie op indirecte antioxidanten zoals SFN, vrije radicalen gegenereerd door de mitochondria [16] of andere fysiologische bronnen van oxidatieve stress [41].
Mitochondria zijn dynamische, subcellulaire organellen die een groot aantal cellulaire functies regelen, gaande van ATP-productie en intracellulaire calciumbuffering tot redoxregulatie en apoptose [13], [49]. Mitochondria vertegenwoordigen ook de belangrijkste bron van reactieve zuurstofspecies (ROS) in de cel. Een goede regulering van de mitochondriale functie is daarom noodzakelijk voor het optimaliseren van de productie van ATP om aan de cellulaire behoeften te voldoen, terwijl tegelijkertijd de potentieel schadelijke effecten van overmatige productie van vrije radicalen worden geminimaliseerd. Een cruciale vereiste voor fijne modulatie van de mitochondriale functie is het vermogen van mitochondriën om zowel onafhankelijk als biochemische machines te functioneren en als onderdeel van een uitgebreid, responsief netwerk.
De morfologie en functie van het mitochondriale netwerk worden bepaald door een gereguleerd evenwicht tussen kernsplijting en fusie. Mitochondriale fissie is vereist voor de overerving van mitochondriën door dochtercellen tijdens celdeling [28] evenals voor de selectieve, autofagische afbraak van gedepolariseerde of beschadigde mitochondriën, genaamd mitofagie [1]. Omgekeerd is fusie vereist voor het complementeren van mitochondriale genomen en het delen van componenten van de elektronentransportketen tussen naburige mitochondria [54]. Op moleculair niveau worden mitochondriale fissie en fusie gereguleerd door grote, dynamineachtige GTPasen. Drie enzymen reguleren voornamelijk fusie: Mitofusins 1 en 2 (Mfn1 / 2) zijn twee passerende buitenmembraaneiwitten die buitenmembraan fusie bemiddelen via heterotypische interacties tussen aangrenzende mitochondria [15], [25], [37], terwijl OPA1 een innerlijke is membraaneiwit dat tegelijkertijd zorgt voor matrixconnectiviteit door het samensmelten van binnenmembranen [5] te reguleren. De GTPase-activiteit van alle drie de eiwitten is vereist voor robuuste fusie [5], [18] en OPA1 wordt verder gereguleerd door complexe proteolyse in het mitochondriale binnenmembraan door de proteasen OMA1 [14], PARL [6] en YME1L [45 ]. Belangrijk is dat intact mitochondriaal membraanpotentieel vereist is voor efficiënte fusie om de integratie van beschadigde en gezonde mitochondria [26] te onderdrukken.
Mitochondriale fissie wordt voornamelijk gekatalyseerd door een cytosolisch eiwit genaamd Dynaminegerelateerd eiwit 1 (Drp1 / DNM1L). Drp1 wordt gerekruteerd uit het cytosol naar mogelijke plaatsen van splijting op het mitochondriale buitenmembraan [43]. De belangrijkste receptoren voor Drp1 op het buitenmembraan zijn mitochondriale fissiefactor (Mff) [32] en, in mindere mate, Fission 1 (Fis1) [51]. Daarnaast werd een lok-receptor, MIEF1 / MiD51, ontdekt die werkt om de activiteit van Drp1-eiwit op potentiële splijtingsites [58] verder te beperken. Eenmaal gekoppeld aan de mitochondriale buitenmembraan, oleriseert Drp1 in spiraalachtige structuren rond het lichaam van de mitochondrion en maakt vervolgens gebruik van de energie die is afgeleid van GTP-hydrolyse om de fysieke splitsing van de mitochondriale buitenste en binnenste membranen [17] te bewerkstelligen. Endoplasmatisch reticulum-afgeleide tubuli werken als een eerste constrictor van mitochondriën voorafgaand aan Drp1 oligomerisatie, wat de openbaring onderstreept dat niet-ingesnoerde mitochondriën breder zijn dan de toegestane omtrek van een voltooide Drp1-spiraal [12]. Actinedynamica is ook belangrijk voor de ER-mitochondria-interacties die voorafgaan aan mitochondriale splijting [24]. Naast zijn rol in mitochondriale fissie katalyseert Drp1 de splijting van peroxisomen [40].
Drp1 lijkt sterk op het goed gekarakteriseerde dynamineproteïne omdat beide eiwitten een N-terminaal GTPase-domein bevatten, een middelste domein dat van cruciaal belang is voor zelfoligomerisatie en een C-terminaal GTPase-effectordomein [31]. Drp1 bereikt selectiviteit voor mitochondriale membranen door een combinatie van interacties met zijn receptoreiwitten Mff en Fis1 en ook door zijn affiniteit voor het mitochondria-specifieke fosfolipide cardiolipine via het unieke B-insert domein van Drp1 [2]. Drp1 bestaat meestal als een homotetrameer in het cytoplasma en assemblage op hogere orde op mitochondriale fissiepunten wordt gemedieerd door het middelste domein van Drp1 [3].
Gezien de impliciete link tussen de mitochondriale functie en de KEAP1-Nrf2-ARE-route, onderzochten we de effecten van Nrf2-activering op de mitochondriale structuur en functie. We demonstreren hier dat SFN mitochondriale hyperfusie induceert die, onverwacht, onafhankelijk is van zowel Nrf2 als KEAP1. Dit effect van SFN is door een remming van de Drp1-functie. We demonstreren verder dat SFN resistentie verleent tegen apoptose die Nrf2-onafhankelijk is en die de waargenomen cellen nabootst die zijn verarmd door Drp1. Deze gegevens geven gezamenlijk aan dat naast het stabiliseren en activeren van Nrf2, SFN de mitochondriale dynamiek moduleert en de cellulaire conditie en overleving behoudt.
Resultaten
Sulforafaan induceert Nrf2/KEAP1-onafhankelijke hyperfusie van mitochondriën
Tijdens het bestuderen van de effecten van Nrf2-activering op de dynamiek van het mitochondriale netwerk, ontdekten we dat behandeling van onsterfelijk gemaakte, menselijke retinale pigmentepitheelcellen (RPE-1) met sulforafaan (SFN), een krachtige activator van Nrf2-signalering, een robuuste fusie veroorzaakte van het mitochondriale netwerk in vergelijking met met drager behandelde controlecellen (Fig. 1A en B). De morfologie van de mitochondriën in deze cellen leek sterk op die van de mitochondriën in cellen die waren uitgeput door siRNA van endogeen Drp1, de belangrijkste mitochondriale splitsingsfactor (Fig. 1A). Dit resultaat bracht het intrigerende idee naar voren dat de mitochondriale splijtings- en fusiestatus direct reageert op Nrf2-niveaus in de cel. Stimulatie van cellen met andere Nrf2-stabilisatoren en activatoren zoals de proteasoomremmer MG132, de pro-oxidant tBHQ of knockdown van de Nrf2-remmer KEAP1 induceerde echter geen mitochondriale fusie (Fig. 1A en B). Stabilisatie van Nrf2 door deze manipulaties werd bevestigd door western blotting voor endogeen Nrf2 (Fig. 1C). Bovendien was expressie van Nrf2 overbodig voor SFN-geïnduceerde mitochondriale fusie, omdat knockdown van endogeen Nrf2 met siRNA dit fenotype niet kon tegengaan (figuur 1D F). Omdat SFN de KEAP1-Nrf2-ARE-route stimuleert door cysteïneresiduen van KEAP1 [21] covalent te modificeren, hebben we KEAP1 uitgeschakeld om te onderzoeken of SFN-geïnduceerde mitochondriale hyperfusie wordt gestimuleerd via een KEAP1-afhankelijke, maar Nrf2-onafhankelijke route. De uitputting van KEAP1 kon echter ook de door SFN geïnduceerde mitochondriale fusie niet opheffen (figuur 1G I). In feite keerde SFN de profissiemorfologie om die werd geïnduceerd door uitputting van KEAP1 (Fig. 1G, paneel b versus paneel d). Deze resultaten geven aan dat SFN-behandeling mitochondriale fusie veroorzaakt, onafhankelijk van de canonieke KEAP1-Nrf2-ARE-route, en leidde ons tot het ondervragen of SFN rechtstreeks invloed heeft op componenten van de mitochondriale splijtings- of fusiemachines.
Figuur 1 SFN induceert NrF2 / KEAP1-onafhankelijke mitochondriale fusie. (A) RPE-1-cellen werden getransfecteerd met de aangegeven siRNA's en 3 dagen later behandeld met DMSO of de Nrf2-activatoren SFN (50 μM), MG132 (10 μM) of tBHQ (100 μM) gedurende 4 uur. Mitochondriën (rood) worden gelabeld met een anti-Tom20-antilichaam en kernen (blauw) worden tegengekleurd met DAPI. (B) grafiek met kwantificering van mitochondriale morfologie scoren van (A). > 50 cellen per aandoening werden op een blinde manier geëvalueerd. (C) vertegenwoordiger western blots van (A). (D) RPE-1-cellen werden getransfecteerd met 10 nM siRNA en 3 dagen later behandeld met SFN gedurende 4 uur voordat ze werden gefixeerd en gekleurd zoals in (A). (E) grafiek met kwantificering van de mitochondriale fenotype-score van (D). > 100 cellen per aandoening werden op een blinde manier geëvalueerd. (F) vertegenwoordiger western blots van (D). (G) Cellen werden getransfecteerd en behandeld zoals in (D) met siCON of siKEAP1. (H) Cellen uit (G) werden gescoord zoals in (B) en (E) op basis van mitochondriale morfologie. (I) Representatieve western blots van (G). Gegevens in (B), (E) en (H) werden samengesteld uit 3 onafhankelijke experimenten elk en de statistische significantie werd bepaald door de tweezijdige Student's t-test. Foutbalken geven +/- SD weer (voor interpretatie van de verwijzingen naar kleur in deze figuurlegenda wordt de lezer verwezen naar de webversie van dit artikel).
Sulforafaan schaadt de mitochondriale vereniging van Drp1
Gebaseerd op de bevinding dat SFN-behandeling mitochondriale hyperfusie induceert, redeneerden we dat dit fenotype een gevolg was van overmatige fusieactiviteit of een remming van fissie-activiteit. Om onderscheid te maken tussen deze twee mogelijkheden, vergeleken we de morfologie van peroxisomen in de aanwezigheid en afwezigheid van SFN. Peroxisomen lijken op mitochondriën omdat ze dynamische organellen zijn waarvan de vorm en lengte constant in beweging zijn [44]. Peroxisomen bevatten zowel Fis1 als Mff in hun buitenmembraan en zijn als gevolg daarvan doelen voor door Drp1 gemedieerde fissie [22], [23]. Peroxisomen maken echter geen gebruik van de fusie-machine van het mitochondriale netwerk en ondergaan daarom geen fusie [39]. Integendeel, peroxisomale fissie wordt tegengewerkt door de verlenging van bestaande peroxisomen via de novo toevoeging van membranen en eiwitten [44]. Omdat peroxisomen geen Mfn1 / 2 en OPA1 hebben, redeneerden we dat als SFN de fusiemachines activeert in plaats van de splijtingsmachines te remmen, de peroxisoomlengte niet zou worden beïnvloed. In met drager behandelde cellen worden peroxisomen gehandhaafd als korte, ronde, puntvormige organellen (Fig. 2, panelen b en d). Echter, SFN-behandeling verhoogde de peroxisoomlengte met ~ 2-vouw in vergelijking met controlecellen (Fig. 2, panelen f en h). Bovendien werden veel van de peroxisomen dicht bij het midden geknepen, wat wijst op een mogelijk scheurdefect (Fig. 2, paneel h, pijlpunten). Evenzo waren peroxisomen in cellen getransfecteerd met Drp1 siRNA abnormaal lang (Fig. 2, panels j en l), wat bevestigt dat Drp1 vereist is voor peroxisomale splijting en suggereert dat SFN-behandeling mitochondriale en peroxisomale fenotypen veroorzaakt door de splijtingsmachinerie te verstoren.
Figuur 2 SFN induceert peroxisomale verlenging. (A) RPE-1-cellen werden getransfecteerd met 10 nM van het aangegeven siRNA en 3 dagen later gedurende 50 uur behandeld met DMSO of 4 µM SFN. Peroxisomen (groen) werden gelabeld met een anti-PMP70-antilichaam, mitochondriën met MitoTracker (rood) en DNA tegengekleurd met DAPI. Vergrote inzet van peroxisomen worden aan de rechterkant getoond (panelen d, h en l) om visualisatie van de veranderingen in morfologie geïnduceerd door SFN- en Drp1-uitputting te vergemakkelijken. Pijlpunten markeren vernauwingspunten. (Voor interpretatie van de verwijzingen naar kleur in deze figuurlegenda wordt de lezer verwezen naar de webversie van dit artikel).
Vervolgens hebben we bepaald hoe SFN de Drp1-functie beperkt. Mogelijkheden waren onder meer verlagingen van expressieniveaus, rekrutering / retentie bij mitochondriën, oligomerisatie of enzymatische activiteit van het GTPase. Een tekort in een van deze zou resulteren in verminderde mitochondriale splijting en hyperfusie. We hebben geen reproduceerbare veranderingen in Drp1-eiwitniveaus gedetecteerd na SFN-behandeling (Fig. 1C en 3A), en concludeerden daarom dat SFN de stabiliteit of expressie van Drp1 niet verandert, consistent met Drp1 met een halfwaardetijd van> 10 uur [50] en onze SFN-behandelingen zijn van kortere duur. Vervolgens hebben we onderzocht of SFN de rekrutering of retentie van Drp1 naar mitochondriën beïnvloedde. Fractioneringsstudies toonden aan dat SFN een verlies van Drp1 uit de mitochondriale fractie induceerde (Fig. 3A, lanen 7-8 en Fig. 3B). Zoals eerder gemeld [43], is slechts een kleine fractie van Drp1 (~ 3%) op elk willekeurig moment geassocieerd met het mitochondriale netwerk tijdens steady-state-omstandigheden, waarbij het grootste deel van het enzym zich in het cytoplasma bevindt (Fig. 3A, banen 5 ). Deze fractioneringsgegevens werden bevestigd met behulp van co-lokalisatie-analyse die een reductie van ~ 8% liet zien in mitochondriën-gelokaliseerde, puntvormige Drp40-foci na SFN-behandeling (Fig. 1C en D). Samen geven deze gegevens aan dat de mitochondriale fusie geïnduceerd door SFN, althans gedeeltelijk, te wijten is aan de verzwakte associatie van Drp3 met de mitochondriën. Onze gegevens maken geen onderscheid tussen of SFN interfereert met de mitochondriale rekrutering versus de mitochondriale retentie van Drp1 of beide, aangezien de analyse van endogene Drp1 niet geschikt was om de GTPase te visualiseren door middel van live-celmicroscopie.
Figuur 3 SFN veroorzaakt een verlies van Drp1 uit de mitochondria. (A) Subcellulaire fractionering van RPE-1-cellen na 4 h van DMSO of SFN. Whole-cellysaten (WCL), nucleaire (Nuc), cytosolische (Cyto) en ruwe mitochondriale (Mito) fracties werden gescheiden door SDS-PAGE en verwerkt voor western blotting met de aangegeven antilichamen. De migratie van molecuulgewichtsmarkers is links aangegeven. (B) Grafieken die densitometrische kwantificatie van Drp1 in de aangegeven fracties van (A) tonen. (C) RPE-1-cellen werden getransfecteerd met 10 nM siCON of siDrp1 en 3 dagen later behandeld met DMSO of SFN voor 4 h. Drp1 (groen) werd gevisualiseerd met een anti-Drp1-antilichaam, mitochondria met MitoTracker (rood) en kernen met DAPI (blauw). (D) Geautomatiseerde co-lokalisatie-analyse van het Drp1- en MitoTracker-signaal van (C). Gegevens in (B) en (D) werden gecompileerd van respectievelijk 3- en 5-onafhankelijke experimenten en de statistische significantie werd bepaald door de tweestaart Student's t-test. Foutbalken geven +/- SD weer en sterretjes duiden statistische significantie aan. (Voor interpretatie van de verwijzingen naar kleuren in deze figuurlegenda wordt de lezer verwezen naar de webversie van dit artikel).
Sulforafaan geeft bescherming tegen door staurosportine geïnduceerde apoptose onafhankelijk van Nrf2
Eerder werk heeft aangetoond dat mitochondriale splijting toelaatbaar is bij de vorming van poriën in het buitenste mitochondriale membraan gegenereerd door Bax/Bak tijdens apoptose [11]. Het is aangetoond dat Drp1 selectief wordt gerekruteerd naar mitochondriën tijdens apoptose [11] en, in overeenstemming hiermee, zijn gefragmenteerde mitochondriën vroeg in het proces waargenomen [27]. Omgekeerd wordt aangenomen dat het remmen van mitochondriale splijting apoptose remt door de vorming van de buitenmembraanporiën te blokkeren die cytochroom c-afgifte mogelijk maken [53]. Dienovereenkomstig vertraagt het stimuleren van mitochondriale fusie de progressie van apoptose die wordt geïnduceerd door verbindingen zoals staurosporine (STS) [14]. Om te bepalen of SFN RPE-1-cellen beschermt tegen STS-gemedieerde apoptose en zo ja, of dit Nrf2 vereist, hebben we een test opgezet om gemakkelijk poly ADP-ribosepolymerase (PARP) splitsing te induceren, een substraat van geactiveerd caspase-3 en definitieve marker van apoptose. Behandeling van RPE-1-cellen met 1 M STS gedurende 6 uur veroorzaakte slechts een zeer bescheiden splitsing van PARP, maar dit werd voorkomen door gelijktijdige SFN-behandeling (bijv. Fig. 4A, baan 3 versus 4). Om de robuustheid van deze test te vergroten, hebben we cellen verder gesensibiliseerd voor STS-geïnduceerde apoptose door ze voor te behandelen met siRNA gericht op de anti-apoptotische factor, Bcl-XL. Deze voorbehandeling verminderde de expressie van Bcl-XL en bevorderde duidelijk PARP-splitsing als een functie van de tijd blootgesteld aan STS (Fig. 4B, vergelijk baan 2 met banen 4-10). Belangrijk is dat 2 uur voorbehandeling met SFN PARP-splitsing in cellen die zijn blootgesteld aan STS verzachtte (Fig. 4C, baan 3 versus 4 en baan 5 versus 6). Evenzo werden cellen die door CRISPR / Cas2 stabiel waren uitgeput van Nrf9 op vergelijkbare wijze beschermd tegen STS-toxiciteit door SFN-voorbehandeling (Fig. 4C, baan 11 versus 12 en baan 13 versus 14 en Fig. 4D). Deze bescherming werd waargenomen met zowel PARP-splitsing (Fig. 4C en D) als cellulaire morfologie (Fig. 4E) als uitlezingen. De werkzaamheid van Nrf2-uitputting door CRISPR / Cas9 werd bevestigd door western-blotting (Fig. 4C, Nrf2-blot). Zoals voorspeld, blokkeerden uitputtende cellen van Drp1, die ook een hyperfusiefenotype opleveren (Fig. 1A), ook PARP-splitsing als reactie op STS in vergelijking met controlecellen die waren geïncubeerd met SFN (Fig. 4F en G). Samen zijn deze bevindingen consistent met SFN dat bescherming biedt tegen apoptose door zijn vermogen om de Drp1-functie te beperken, onafhankelijk van de stabilisatie en activering van Nrf2.
Figuur 4 De cytoprotectieve effecten van SFN zijn onafhankelijk van Nrf2-expressie (A) RPE-1-cellen werden voorbehandeld met DMSO of 50 µM SFN gedurende 2 uur voorafgaand aan behandeling met DMSO, 1 µM staurosporine (STS) of 50 µm SFN. M etoposide gedurende 6 uur en werden verwerkt voor anti-PARP western blotting. (B) RPE-1-cellen werden getransfecteerd met 2.5 nM siCON, 1 nM siBcl-XL of 2.5 nM siBcl-XL en 3 dagen later werden ze behandeld met DMSO of 1 µM STS gedurende 2, 4 of 6 uur. Representatieve Western-blots worden getoond en de migratie van molecuulgewichtmarkers wordt aan de linkerkant aangegeven. (C) CRISPR / Cas9-gegenereerde wildtype (Nrf2WT) en Nrf2 knock-out (Nrf2KO) RPE-1-cellen werden getransfecteerd met 1 nM siBcl-XL en 3 dagen later werden ze voorbehandeld met DMSO of 50 μM SFN gedurende 2 uur . Vervolgens werden de cellen gedurende 1, 2 of 4 uur behandeld met 6 µM STS. Representatieve Western-blots met de aangegeven antilichamen worden weergegeven. (D) Kwantificering van gesplitst PARP als een percentage van totaal PARP (gesplitst + ongesplitst) uit 3 onafhankelijke experimenten. Belangrijk is dat de niveaus van gesplitst PARP vergelijkbaar waren, ongeacht of cellen Nrf2 tot expressie brachten of niet, wat aangeeft dat SFN-bescherming tegen STS onafhankelijk is van de transcriptiefactor. (E) 20X fasecontrastbeelden die onmiddellijk voorafgaand aan de oogst van lysaten van (C) zijn genomen. Schaalbalk = 65 m. (F) Representatieve western blots die aantonen dat uitputting van Drp1 een bijna vergelijkbare bescherming biedt tegen STS als SFN-behandeling. RPE-1-cellen werden getransfecteerd met 1 nM siBcl-XL en bovendien getransfecteerd met ofwel 10 nM siCON of 10 nM siDrp1. 3 dagen later werden siCON-cellen voorbehandeld met SFN zoals in (A) en (C) en vervolgens 4 uur blootgesteld aan STS voordat ze werden geoogst en verwerkt voor western blotting met de aangegeven antilichamen. (G) Hetzelfde als (D) voor de gegevens gepresenteerd in (F) samengesteld uit 3 onafhankelijke experimenten. Foutbalken weerspiegelen +/- SEM
Discussie
We hebben ontdekt dat SFN mitochondriale fissie / fusiedynamiek moduleert, onafhankelijk van de effecten op de KEAP1-Nrf2-ARE-route. Dit is intrigerend vanwege een veronderstelde koppeling tussen mitochondriale dysfunctie en ROS-productie en de noodzaak om squelching mitochondria-afkomstige vrije radicalen door de activering van Nrf2. Deze extra functionele impact van SFN is van potentieel belang gezien de meer dan 30 klinische trials die momenteel aan de gang zijn en SFN testen voor de behandeling van een verscheidenheid aan ziekten, waaronder prostaatkanker, obstructieve longziekte en sikkelcelziekte [7], [10], [ 47].
Omdat SFN een isothiocyanaat is [56] en het Nrf2-signalering activeert door direct kritische KEAP1-cysteïnes te acyleren om Nrf2-degradatie [21] te onderdrukken, volgt hieruit dat SFN zijn pro-fusie-effecten uitoefent door de activiteit van een splijtings- of fusiefactor te moduleren via cysteïnemodificatie . Onze gegevens ondersteunen sterk dat Drp1 negatief wordt gereguleerd door SFN, hoewel het nog moet worden opgehelderd of de GTPase een direct doelwit van acylering is. Ondanks deze kenniskloof wordt de functie van Drp1 duidelijk aangetast door SFN, aangezien zowel mitochondriën als peroxisomen hyperfuseren als reactie op SFN-behandeling en deze organellen Drp1 delen voor hun respectievelijke splitsingsgebeurtenissen [38]. Bovendien vermindert SFN de hoeveelheid Drp1 die lokaliseert en zich ophoopt bij mitochondriën (Fig. 3). Omdat onze experimenten werden gedaan met alle endogene eiwitten, is onze detectie van Drp1 op mitochondriale splijtingsplaatsen onder stabiele omstandigheden, en bijgevolg kunnen we geen onderscheid maken tussen een rekruterings- versus een retentiedefect van het enzym veroorzaakt door SFN. Verder kunnen we de mogelijkheid niet uitsluiten dat SFN een receptor in de mitochondriën (Fis1 of Mff) acyleert om Drp1-rekrutering te blokkeren, we vermoeden dat Drp1 direct is gemodificeerd. Drp1 heeft negen cysteïnes, waarvan er acht zich bevinden in het middelste domein dat nodig is voor oligomerisatie [3], en waarvan er één zich bevindt in het GTPase Effector Domain (GED) aan de C-terminus van Drp1. Directe acylering van een van deze cysteïnes zou een activiteitsdefect in Drp1 kunnen veroorzaken en daarom ten grondslag liggen aan het effect van SFN op de mitochondriale dynamiek. Met name suggereert eerder werk dat defecten in oligomerisatie en katalytische activiteit de retentie van Drp1 in de mitochondriën kunnen tenietdoen [52]. Cys644 in het GED-domein is een bijzonder aantrekkelijk doelwit op basis van eerder werk dat aantoont dat mutatie van deze cysteïne mutaties kopieert die de Drp1-GTPase-activiteit [4] aantasten en dat deze specifieke cysteïne wordt gemodificeerd door thiol-reactieve elektrofielen [9]. Om deze openstaande vraag op te lossen, is massaspectrometrische validatie vereist. Samenvattend hebben we een nieuwe, cytoprotectieve functie geïdentificeerd voor de klinisch relevante verbinding SFN. Naast het activeren van de belangrijkste anti-oxidant transcriptiefactor Nrf2, bevordert SFN mitochondriale en peroxisomale fusie, en dit effect is onafhankelijk van Nrf2. Het mechanisme dat aan dit fenomeen ten grondslag ligt, omvat een vermindering van de functie van de GTPase Drp1, de primaire bemiddelaar van mitochondriale en peroxisomale splitsing. Een belangrijk gevolg van SFN-gemedieerde mitochondriale fusie is dat cellen resistent worden tegen de toxische effecten van de apoptose-inductor staurosporine. Deze aanvullende cytoprotectieve werking van SFN zou van bijzonder klinisch nut kunnen zijn bij de talrijke neurodegeneratieve ziekten waarvoor leeftijd de belangrijkste risicofactor is (bijv. de ziekte van Parkinson, de ziekte van Alzheimer, leeftijdsgebonden maculaire degeneratie), aangezien deze ziekten in verband zijn gebracht met apoptose en verminderd niveaus en / of ontregeling van Nrf2 [35], [36], [48].
Materialen en methoden
Apoptosis-assays
Cellen werden gezaaid en getransfecteerd met siRNA zoals hieronder aangegeven. De cellen werden gedurende 50 uur voorbehandeld met 2 µM sulforafaan om mitochondriale fusie te induceren en werden vervolgens behandeld met 1 µM staurosporine om apoptose te induceren. Op het moment van de oogst werden de media verzameld in afzonderlijke buisjes en onderworpen aan centrifugatie met hoge snelheid om apoptotische cellen te pelleteren. Deze celpellet werd gecombineerd met hechtende cellen en opgelost in 2 keer geconcentreerde Laemmli-buffer. Monsters werden onderworpen aan anti-PARP western blotting.
CRISPR / Cas9 Construct-generatie
Om LentiCRISPR / eCas9 1.1 te maken, werd LentiCRISPR v2 (addgene #52961) voor het eerst geknipt met Age1 en BamH1. Vervolgens werd SpCas9 van eSpCas9 1.1 (addegen # 71814) met PCR geamplificeerd met Age1 en BamH1 overhangen met behulp van de volgende primers (Forward AGCGCACCGGTTCTAGAGCGCTGCCACCATGGACTATAAGGACCACGAC, Reverse AAGCGCGGATCCCTTTTTCTTTTTTGCCTGGCCGG) en geligeerd in de snijvector hierboven. sgRNA-sequenties werden bepaald door Benchling.com te gebruiken. Parameters werden ingesteld om de coderende sequentie te targeten met de hoogste on-target en laagste off-target scores. De volgende sequenties (doelsequentie onderstreept hs sgNFE2L2 # 1 sense CACCGCGACGGAAAGAGTATGAGC, antisense AAACGCTCATACTCTTTCCGTCGC; hs sgNFE2L2 # 2 sense CACCGGTTTCTGACTGGATGTGCT, antisense AAACAGCACATCCAGTCAGAAACC; hs sgNFE2L2 # 3 sense CACCGGAGTAGTTGGCAGATCCAC, antisense AAACGTGGATCTGCCAACTACTCC) werden gekoppeld en geligeerd in BsmB1 gesneden LentiCRISPR / eCas9 1.1. Lentiviraal geïnfecteerde RPE-1-cellen werden geselecteerd met puromycine en gehandhaafd als een gepoolde populatie. Knockout werd bevestigd door immunofluorescentie en Western-blotting.
Celkweek en transfecties
Menselijke retinale pigmentepitheelcellen getransformeerd met telomerase (RPE-1) (ATCC) werden gekweekt in Dulbecco's Modified Eagle Medium (DMEM) met 1 g / l glucose aangevuld met penicilline, streptomycine, 1x niet-essentiële aminozuurcocktail (Life Technologies), en 10% foetaal runderserum (Life Technologies). Voor siRNA-transfecties werden 30,000-35,000 cellen / ml gedurende de nacht gezaaid. Cellen ontvingen 10 nM siRNA verdund in serumvrij DMEM en gecombineerd met 0.3% interferine-transfectiereagens (PolyPlus). Voor sensibilisatie van apoptose ontvingen cellen 1 nM Bcl-XL-siRNA. Cellen werden 2-3 dagen na transfectie geoogst.
Chemicaliën, antilichamen en siRNA Oligos
Antilichamen tegen β-tubuline (celsignalering), β-tubuline (Sigma), Drp1 (BD Biosciences), KEAP1 (Proteintech), Lamin B1 (Abcam), PARP (Cell Signaling), PMP70 (Abcam) en Tom20 (BD Biosciences) ) werden gebruikt in verdunningen van 1: 1000 voor western blotting en voor immunofluorescentie. Intern werd anti-Nrf2 konijnenantilichaam gebruikt in 1: 2000 voor western blotting [34], [59]. Sulforafaan (Sigma) en staurosporine (Tocris) werden gebruikt bij respectievelijk 50 µM en 1 µM. siRNA's tegen Drp1 (Dharmacon), Nrf2 (Dharmacon), KEAP1 (celsignalering) en Bcl-XL (celsignalering) werden gebruikt bij 10 nM tenzij anders vermeld.
Immunofluorescentie en in Vivo Labelling
Cellen gezaaid op glazen dekglaasjes van 18 mm werden behandeld met drager of medicijn, gefixeerd in 3.7% formaldehyde en vervolgens gedurende 0.2 minuten gepermeabiliseerd in 100% Triton X-10 / PBS op ijs. Primaire antilichamen werden een nacht bij 3 ° C geïncubeerd in 4% runderserumalbumine (BSA) in PBS. Na PBS-wasbeurten werden de cellen gedurende 1 uur geïncubeerd in soortgeschikte, Alexa488- of Alexa546-, geconjugeerde secundaire antilichamen (verdund 1: 1000) en 0.1 µg / ml DAPI (Sigma) in 3% BSA / PBS. Mitochondria werden zichtbaar gemaakt door ofwel anti-Tom20 immunofluorescentie ofwel door cellen te incuberen in 200 nM MitoTracker Red CMXRos (Molecular Probes, Inc.) in serumvrij DMEM gedurende 30 minuten bij 37 ° C voorafgaand aan fixatie.
Microscopie en beeldanalyse
Immunofluorescentiemonsters werden bekeken op een LSM710 confocale microscoop (Carl Zeiss). Microfoto's werden gemaakt met behulp van 63X of 100X olie-immersiedoelstellingen en afbeeldingen aangepast en verbeterd met Adobe Photoshop CS6. Co-lokalisatie-analyse werd uitgevoerd met behulp van de Carl Zeiss LSM710-co-lokalisatiefunctie met handmatig ingestelde drempels terwijl men blind was voor de identiteit van de monsters. Schaalbalken overal, tenzij anders aangegeven, zijn 10 m. Mitochondriale morfologie werd beoordeeld door middel van geblindeerde scores. Als de mitochondriën van een cel werden gehandhaafd als meerdere, ronde, onderscheiden puncta, werd de cel gescoord als splitsing . Als individuele mitochondriën niet te onderscheiden waren en het hele mitochondriale netwerk continu leek, werd de cel gescoord als fusie . Alle andere cellen, inclusief die met clusterende mitochondriën, werden gescoord als intermediair .
Subcellulaire fracties
RPE-1-cellen werden tot confluentie gekweekt. Na een PBS-wassing werden de cellen gedurende 600 minuten bij 10 gg gecentrifugeerd en opnieuw gesuspendeerd in 600 µl isolatiebuffer (210 mM mannitol, 70 mM sucrose, 5 mM MOPS, 1 mM EDTA pH 7.4 + 1 mM PMSF). De suspensie werd 30 keer gelyseerd in een Dounce-homogenisator. Een fractie van het homogenaat werd geconserveerd als een heel cellysaat De rest werd gedurende 800 minuten bij 10 g gecentrifugeerd om kernen te pelleteren. Supernatanten werden gedurende 1500 minuten bij 10 g gecentrifugeerd om de resterende kernen en niet-gelyseerde cellen te verwijderen. Dit supernatant werd gedurende 15,000 minuten bij 15 g gecentrifugeerd om mitochondriën te pelleteren. Het supernatant werd bewaard als de cytosolische fractie . De pellet werd voorzichtig gewassen met PBS en opnieuw gesuspendeerd in isolatiebuffer. De eiwitconcentratie van elke fractie werd gemeten met bicinchoninezuur (BCA) -test en equivalente hoeveelheden eiwit werden gescheiden door SDS-PAGE.
Western Blotting
Cellen werden gewassen in PBS en opgelost in 2 keer geconcentreerde Laemmli solubiliserende buffer (100 mM Tris [pH 6.8], 2% SDS, 0.008% broomfenolblauw, 2% 2-mercapto-ethanol, 26.3% glycerol en 0.001% Pyrinine Y). Lysaten werden 5 minuten gekookt voorafgaand aan het laden op natriumdodecylsulfaat (SDS) polyacrylamidegels. Eiwitten werden overgebracht naar nitrocellulosemembranen en de membranen werden gedurende 1 uur geblokkeerd in 5% melk/TBST. Primaire antilichamen werden verdund in 5% melk/TBST en een nacht bij 4 °C met de blot geïncubeerd. Met mierikswortelperoxidase (HRP) geconjugeerde secundaire antilichamen werden verdund in 5% melk/TBST. Blots werden verwerkt met verbeterde chemiluminescentie en densitometrische kwantificaties werden uitgevoerd met behulp van ImageJ-software.
Sulforafaan is een chemische stof uit de isothiocyanaatcollectie van organozwavelverbindingen verkregen uit kruisbloemige groenten, waaronder broccoli, kool, bloemkool, boerenkool en collards, onder andere. Sulforafaan wordt geproduceerd wanneer het enzym myrosinase glucoraphanine, een glucosinolaat, omzet in sulforafaan, ook bekend als sulforafaan-glucosinolaat. Broccolispruiten en bloemkool hebben de hoogste concentratie glucoraphanine of de voorloper van sulforafaan. Onderzoeksstudies hebben aangetoond dat sulforafaan de antioxidantvermogens van het menselijk lichaam verbetert om verschillende gezondheidsproblemen te voorkomen. Dr. Alex Jimenez DC, CCST Insight
Sulforafaan en de effecten ervan op kanker, mortaliteit, veroudering, hersenen en gedrag, hartaandoeningen en meer
Isothiocyanaten zijn enkele van de belangrijkste plantaardige stoffen die u in uw dieet kunt krijgen. In deze video maak ik de meest uitgebreide case voor hen die ooit is gemaakt. Korte aandachtsspanne? Ga naar uw favoriete onderwerp door op een van de onderstaande tijdspunten te klikken. Volledige tijdlijn hieronder.
Belangrijkste secties:
00: 01: 14 - Kanker en sterfte
00: 19: 04 - Veroudering
00: 26: 30 - Hersenen en gedrag
00: 38: 06 - Laatste overzicht
00: 40: 27 - Dosis
Volledige tijdlijn:
00: 00: 34 - Introductie van sulforafaan, een belangrijk aandachtspunt van de video.
00: 01: 14 - Kruisbloemige groenteconsumptie en verlagingen van mortaliteit door alle oorzaken.
00: 02: 12 - Risico op prostaatkanker.
00: 02: 23 - Blaaskankerrisico.
00: 02: 34 - Longkanker bij rokers risico.
00: 02: 48 - Risico op borstkanker.
00: 03: 13 - Hypothetisch: wat als je al kanker hebt? (Interventie)
00: 03: 35 - Aannemelijk mechanisme dat de associatieve gegevens over kanker en mortaliteit stuurt.
00: 04: 38 - Sulforafaan en kanker.
00: 05: 32 - Dierlijk bewijs dat een sterk effect van extract van broccolispruit toont op de ontwikkeling van blaastumoren bij ratten.
00: 06: 06 - Effect van directe suppletie van sulforafaan bij prostaatkankerpatiënten.
00: 07: 09 - Bioaccumulatie van isothiocyanaatmetabolieten in daadwerkelijk borstweefsel.
00: 08: 32 - Remming van stamcellen van borstkanker.
00: 08: 53 - Geschiedenisles: brassica's werden vastgesteld met gezondheidseigenschappen zelfs in het oude Rome.
00: 09: 16 - het vermogen van Sulforaphane om de kankerverwekkende uitscheiding te verbeteren (benzeen, acroleïne).
00: 09: 51 - NRF2 als een genetische switch via antioxidantresponselementen.
00: 10: 10 - Hoe NRF2-activering de carcinogene excretie via glutathion-S-conjugaten verbetert.
00: 10: 34 - Spruitjes verhogen glutathion-S-transferase en verminderen DNA-schade.
00: 11: 20 - Broccolispruitendrank verhoogt de benzeenuitscheiding met 61%.
00: 13: 31 - Broccoli-spruithomogenaat verhoogt antioxiderende enzymen in de bovenste luchtwegen.
00: 15: 45 - Kruisbloemige groenteconsumptie en sterfte aan hart- en vaatziekten.
00: 16: 55 - Broccolispruitpoeder verbetert de bloedlipiden en het algemene risico op hartziekten bij type 2 diabetici.
00: 19: 04 - Begin van het verouderingsgedeelte.
00: 19: 21 - Met Sulforaphane verrijkte voeding verlengt de levensduur van kevers van 15 tot 30% (onder bepaalde omstandigheden).
00: 20: 34 - Belang van lage ontsteking voor een lang leven.
00: 22: 05 - Kruisbloemige groenten en broccoli-kiempoeder lijken een breed scala aan ontstekingsmarkers bij mensen te verminderen.
00: 24: 14 - Muisstudies suggereren dat sulforaphane de adaptieve immuunfunctie op oudere leeftijd zou kunnen verbeteren.
00: 25: 18 - Sulforafaan verbeterde de haargroei in een muismodel van kalend. Afbeelding bij 00: 26: 10.
00: 26: 30 - Begin van de sectie hersenen en gedrag.
00: 27: 18 - Effect van extract van broccolispruiten op autisme.
00: 27: 48 - Effect van glucoraphanine op schizofrenie.
00: 28: 17 - Begin van depressie discussie (plausibel mechanisme en studies).
00: 31: 21 - Muisstudie met behulp van 10 verschillende modellen van stress-geïnduceerde depressie laten sulforafaan zien, even effectief als fluoxetine (prozac).
00: 32: 00 - Onderzoek toont aan dat directe inname van glucoraphanine bij muizen even effectief is bij het voorkomen van depressies als gevolg van een sociaal nederlaagstressmodel.
00: 33: 01 - Begin van de sectie neurodegeneratie.
00: 33: 30 - Sulforafane en de ziekte van Alzheimer.
00: 33: 44 - Sulforafane en de ziekte van Parkinson.
00: 33: 51 - De ziekte van Sulforaphane en Hungtington.
00: 34: 43 - Begin van een traumatisch gedeelte over hersenletsel.
00: 35: 01 - Sulforafaan wordt geïnjecteerd onmiddellijk nadat TBI het geheugen verbetert (muisstudie).
00: 35: 55 - Sulforafaan en neuronale plasticiteit.
00: 36: 32 - Sulforafaan verbetert het leren in het model van type II diabetes bij muizen.
00: 37: 19 - Sulforafaan en duchenne spierdystrofie.
00: 37: 44 - Myostatinremming in spieratlanticellen (in vitro).
00: 38: 06 - Late video recapitulatie: sterfte en kanker, DNA-schade, oxidatieve stress en ontsteking, uitscheiding van benzeen, cardiovasculaire aandoeningen, type II diabetes, effecten op de hersenen (depressie, autisme, schizofrenie, neurodegeneratie), NRF2-route.
00: 40: 27 - Gedachten over het uitzoeken van een dosis broccoli of sulforafaan.
00: 41: 01 - Anekdotes over ontspruiten thuis.
00: 43: 14 - Op kooktemperaturen en sulforafaanactiviteit.
00: 43: 45 - Darmbacterieomzetting van sulforafaan uit glucoraphanine.
00: 44: 24 - Supplementen werken beter in combinatie met actieve myrosinase van groenten.
00: 44: 56 - Koken technieken en kruisbloemige groenten.
Verhitting vermindert Epithiospecifier Protein Activity en verhoogt Sulforaphane Formation in Broccoli
Abstract
Sulforafaan, een isothiocyanaat uit broccoli, is een van de krachtigste van voedsel afgeleide anticarcinogenen. Deze verbinding is niet aanwezig in de intacte groente, maar wordt gevormd uit zijn glucosinolaatvoorloper, glucoraphanine, door de werking van myrosinase, een thioglucosidase-enzym, wanneer broccoliweefsel wordt fijngemaakt of gekauwd. Een aantal onderzoeken heeft echter aangetoond dat de sulforafaanopbrengst van glucoraphanine laag is, en dat een niet-bioactief nitrilanalogon, sulforafaannitril, het primaire hydrolyseproduct is wanneer plantenweefsel bij kamertemperatuur wordt vermalen. Recent bewijs suggereert dat in Arabidopsis de vorming van nitril uit glucosinolaten wordt gecontroleerd door een warmtegevoelig eiwit, epithiospecifier-eiwit (ESP), een niet-katalytische cofactor van myrosinase. Onze doelstellingen waren om de effecten te onderzoeken van het verwarmen van broccoliroosjes en spruiten op de vorming van sulforafaan en sulforafaannitril, om te bepalen of broccoli ESP-activiteit bevat, en om vervolgens warmteafhankelijke veranderingen in ESP-activiteit, sulforafaangehalte en bioactiviteit te correleren, zoals gemeten door inductie van de fase II ontgiftingsenzym chinonreductase (QR) in celcultuur. Het verwarmen van verse broccoliroosjes of broccolispruiten tot 60°C voorafgaand aan homogenisatie verhoogde tegelijkertijd de vorming van sulforafaan en verminderde vorming van sulforafaannitril. Een significant verlies van ESP-activiteit liep parallel met de afname van de vorming van sulforafaannitril. Verhitten tot 70 �C en hoger verminderde de vorming van beide producten in broccoliroosjes, maar niet in broccolispruiten. De inductie van QR in gekweekte muizenhepatoom Hepa lclc7-cellen gingen gepaard met een toename van de vorming van sulforafaan.
Voorverwarmen van broccoliroosjes en spruiten tot 60 C verhoogde de door myrosinase gekatalyseerde vorming van sulforafaan (SF) in plantaardige weefselextracten na het fijnmaken aanzienlijk. Dit ging gepaard met een afname van de vorming van sulforafaan nitril (SF Nitril) en de activiteit van epithiospecifier proteïne (ESP).
Kortom, sulforafaan is een fytochemische stof die wordt aangetroffen in broccoli en andere kruisbloemige groenten. Een ongecontroleerde hoeveelheid oxidanten, veroorzaakt door zowel interne als externe factoren, kan oxidatieve stress in het menselijk lichaam veroorzaken, wat uiteindelijk kan leiden tot een verscheidenheid aan gezondheidsproblemen. Sulforafaan kan de productie van Nrf2 activeren, een transcriptiefactor die helpt bij het reguleren van beschermende antioxidantmechanismen die de reactie van de cel op oxidanten regelen. De reikwijdte van onze informatie is beperkt tot chiropractische en spinale gezondheidsproblemen. Als u het onderwerp wilt bespreken, kunt u het aan Dr. Jimenez vragen of contact met ons opnemen via�915-850-0900 .
Rugpijn is een van de meest voorkomende oorzaken van arbeidsongeschiktheid en gemiste werkdagen wereldwijd. Rugpijn is de tweede meest voorkomende reden voor doktersbezoeken, alleen overtroffen door bovenste luchtweginfecties. Ongeveer 80 procent van de bevolking zal gedurende zijn leven minstens één keer rugpijn ervaren. De wervelkolom is een complexe structuur die bestaat uit botten, gewrichten, ligamenten en spieren, naast andere zachte weefsels. Hierdoor kunnen verwondingen en / of verergerde omstandigheden, zoals herniated discs, kan uiteindelijk leiden tot symptomen van rugpijn. Sportblessures of ongevallen met auto-ongelukken zijn vaak de meest voorkomende oorzaak van rugpijn, maar soms kunnen de eenvoudigste bewegingen pijnlijke resultaten hebben. Gelukkig kunnen alternatieve behandelingsopties, zoals chiropractische zorg, de rugpijn verlichten door het gebruik van spinale aanpassingen en handmatige manipulaties, waardoor uiteindelijk de pijnverlichting wordt verbeterd.
IFM's Find A Practitioner-tool is het grootste verwijzingsnetwerk in Functional Medicine, opgericht om patiënten te helpen bij het vinden van Functional Medicine-beoefenaars overal ter wereld. IFM Certified Practitioners worden als eerste vermeld in de zoekresultaten, gezien hun uitgebreide opleiding in functionele geneeskunde